Introduction
Panic disorder (PD) is an anxiety disorder characterized by spontaneous and recurrent panic attacks which display multiple physical symptoms [1]. Lifetime prevalence of PD can vary from 0.5 to 4%, with peak age of onset being 25 years. PD is more commonly encountered in some groups, such as females, the unemployed, divorced/separated/widowed, individuals with lower education, and those with low household income [2]. The treatment approach involves psychopharmacological interventions and cognitive behavioral therapy (CBT) or a combination of both [1]. Diagnostic delay may lead to morbidity in psychiatric disorders [3]. A meta-analysis study reported that subjects with PD are nearly 4 times as likely to attempt suicide as compared to subjects without PD [4]. Though the studies up to now have revealed some of the genetic factors, the pathophysiology of the disorder seems to have a complicated genetic background covering the interaction of both genetic and environmental factors [5].
The evolutionary conserved neuropeptide oxytocin (OXT) is involved in many physiological and behavioral mechanisms. Apart from its well-established role in pair-bonding and reproductive behaviors, it has been shown to play significant roles in emotional stability, empathy, trust, mood, social cognition, and affiliation behavior [6–8]. Moreover, in the last few years, it has also drawn attention as a cardiovascular hormone [9]. OXT regulates neurotransmitter and emotional mechanisms via oxytocin receptors (OXTRs), which are expressed in distinct brain regions associated with anxiety and fear such as hypothalamus, amygdala and hippocampus, and on target neuronal cells [7, 10].
The comprehensive evaluation of the role of both oxytocin and its receptor may pave the way for innovative psychiatric approaches in future. The application of intranasal oxytocin seems to be promising in both animal [11, 12] and human clinical studies in terms of anxiety and aggressive behavior management [13, 14].
The OXTR gene, located on chromosome 3p25, consists of four exons and three introns. It encodes a 389-aa polypeptide with seven transmembrane domains belonging to the class I G protein-coupled receptor family [15]. Since the physiologic effects of oxytocin are mediated through its specific receptor (OXTR), both are involved in the regulation of behavioral patterns such as trust, empathy, altruism, affiliation behavior, and stress responsiveness [7, 16]. OXTR variants have been found to be associated with outcomes at different levels in various disorders including aggression [7], depression and separation anxiety [16, 17], autistic disorders [6], alcohol withdrawal symptoms [10], schizophrenia [18], post-traumatic stress disorder [19, 20], and social cognition in bipolar disorder type I [21].
The primary purpose of this study was to investigate the relationships between OXTR gene variants and panic disorder liability. We also evaluated the Separation Anxiety Symptom Inventory (SASI) and the Adult Separation Anxiety Questionnaire (ASA) in panic disorder patients and compared them with variations of the OXTR gene as the second aim. To the best of our knowledge, this is one of the most comprehensive models conducted in panic disorder research, evaluating the effects of OXTR variants in terms of both disease liability and childhood and adult separation anxiety levels.
Material and methods
Participants
In the study, 134 PD patients (age range: 18–71 years mean: 36.34 ±11.53; gender: 59.0% female and 41% male) who met the criteria according to the Diagnostic and Statistical Manual of Mental Disorders, 5th edition were recruited from the Department of Psychiatry, Faculty of Medicine, Ondokuz Mayıs University, Samsun, Turkey. University staff (n = 140) without a personal/family history of psychiatric disorders served as controls (mean age: 39.14 ±9.50; gender: 60.71% female and 39.29% male). The study was approved by the Medical Ethics Committee of Ondokuz Mayıs University (OMU KAEK 2020/709) and complied with the Declaration of Helsinki.
Separation anxiety evaluations
Two validated interviewer-administered instruments were used: SASI [22] and ASA [23]. Turkish versions of the instruments were proven to be reliable and valid [24]. The cut-off values used for SASI and ASA evaluations were ≥ 12 and ≥ 25, respectively.
Genotyping analyses
Peripheral blood samples were collected in EDTA containing tubes and DNA extractions were performed with PureLink Genomic DNA Mini Kit (Invitrogen-Thermo Fisher Scientific, MA, USA). Genotyping analyses were carried out with PCR-RFLP method previously reported in the literature for OXTR rs53576 [16], OXTR rs237902 [6], and OXTR rs2254298 [16], with modifications. PCR reaction was carried out in a 25 µl mixture containing 1 × Taq buffer, 2 mM MgCl2, 0.2 mM dNTP, 0.5 µM of forward and reverse primer pair, 1.5 U Taq DNA polymerase (Thermo Fisher Scientific, MA, USA), and ~2 00 ng of DNA sample. Thermal cycler conditions for OXTR rs53576 were as follows: 35 cycles of 40 s at 94°C, annealing for 45 s at 58°C and extension for 45 s at 72°C. A predenaturation step for 6 min at 94°C and a final extension step for 12 min at 72°C were included. The same thermal cycler conditions were used for OXTR rs2254298 with a difference in annealing temperature (60.5°C). Thermal cycler conditions for OXTR rs237902 consisted of a pre-denaturation step at 95°C for 10 min, 35 cycles of denaturation at 95°C for 45 s, annealing at 62.3°C for 45 s and extension at 72°C for 45 s. A final extension step at 72°C for 12 min completed the reaction. The expected amplicon sizes of 340 bp, 527 bp, and 307 bp for OXTR rs53576, OXTR rs237902, and OXTR rs2254298, respectively, were confirmed. For the analysis of the OXTR rs53576 genotypes, amplified fragments were digested with BamHI restriction enzyme (Thermo Fisher Scientific, MA, USA) at 37°C for 2 h and were separated by electrophoresis. The unrestricted PCR product (GG genotype) had a size of 340 bp, complete restriction (AA genotype) produced bands of 120 bp and 220 bp, and heterozygotes had all the bands. For the analysis of the OXTR rs237902 genotypes, amplicons were digested with Tsp509l restriction enzyme (Thermo Fisher Scientific, MA, USA) at 65°C for 2 h. The complete restriction (AA genotype) resulted in the production of 321 bp and 206 bp fragments, while the GG genotype remained unrestricted (527 bp). For the analysis of OXTR rs2254298, amplicons were digested with Bsrl restriction enzyme (Thermo Fisher Scientific, MA, USA) at 65°C for 2 h. The expected fragment sizes for the analyzed genotypes were: GG: 163 + 101 + 34 + 9 bp and AA: 163 + 135 + 9 bp. A minor set of all the samples (20%) was randomly re-genotyped for the analyzed variants to ensure genotyping success, and the results were found concordant.
Statistical analysis
In order to test the sample size with power analysis, Python programming language and its statsmodels module were used. Python language was chosen because of its strong scientific capabilities, open source code and very good third party module support [25]. The statsmodels module was chosen for its ability to perform power analysis and determine the minimum limit of sample size. This module is known as the gold standard for making reliable statistical calculations [26]. All necessary visualizations were made with the matplotlib module [27]. For sample size determination, the effect size was chosen as 0.25, indicating a small-medium effect size. According to Cohen’s guidelines, this size is often observed in fields such as education, psychology, and social sciences. α = 0.05 was chosen as the classical significance level, which accepts a type I error risk of 5% in hypothesis testing. The target power was determined as 80% in this study. This ensures that when a real effect exists, the probability of correctly detecting this effect is at least 80% [28]. Based on these data, the necessary calculations were made with the Python script developed, and the minimum sample size was found to be 128; it was interpreted that samples of 128 and above were sufficient to support the study’s objectives. The sample size power interaction graph was created with the values given below. At this point, the power target of 80% was achieved with a sample size of n = 128.
R statistical software (version 4.2.2) was used. Using the HWE.chisq subroutine of the genetics package in R, each of the variants was found compatible with Hardy-Weinberg equilibrium (HWE). Pearson’s χ2 test with Yates’ continuity correction and Fisher’s exact test were used to evaluate the associations of the OXTR genotypes/alleles with PD. The statistical significance level was defined as p < 0.05. Odds ratios (ORs) and 95% confidence intervals (CIs) were used to interpret the findings. Using simple Bayesian logistic regression models for SASI and ASA, we investigated the association between the genotypes of OXTR variants and separation anxiety.
Results
Panic disorder data
Nocturnal panic attacks were observed in 41.8% of the patients. A triggering life event was present in 66.5% (death of a close person: 23.60%, life-threatening event: 38.20%, psychosocial stress factor: 38.20%) of the patients. Object carrying was present in 45.5%. Nearly half of the patients had a family history of psychiatric disease (panic disorder: 33.6%, others: 12.7%). Mean age of disease onset was 27.49 ±10.38. Mean age of treatment onset was 30.25 ±10.38.
Case-control study: relationship between OXTR genotypes and panic disorder
The AA genotype and (A allele) were taken as the reference categories. Based on the results of Pearson’s χ2 test with the Yates’ continuity correction, no significant relationship was detected between the genotypes and panic disorder. For OXTR rs237902, the allele frequency appeared to have a significant impact on panic disorder (χ2 = 6.328, p = 0.012). When compared to the G allele frequency, the A allele frequency increases the probability of panic disorder by 1.585 times (Figure 1 A, Table I).
Table I
Genotype and allele comparisons for healthy controls and panic disorder patients
Figure 1
A – Box plots comparing the probabilities of observing healthy controls and panic disorder patients with respect to the OXTR genotypes. B – ANOVA plots comparing the mean age of disease onset with respect to the genotypes of each OXTR gene variant
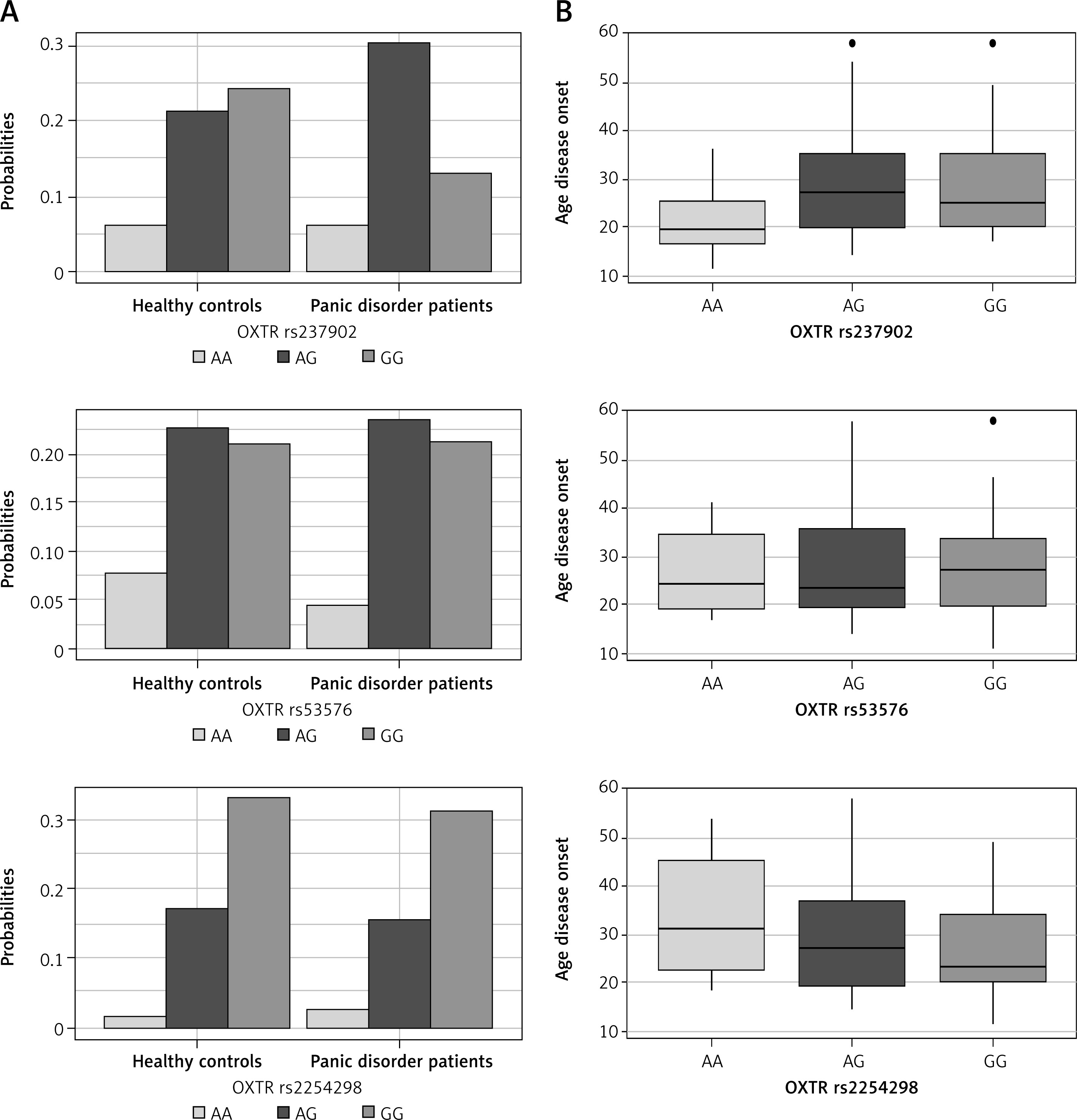
Based on the Fisher’s exact test results, no significant relationship was observed between the levels of the categorical variables and OXTR gene variants (p > 0.05). The analysis of variance (ANOVA) results indicate a difference between the OXTR rs237902 genotypes in terms of the average age of disease onset (F = 3.218, p = 0.043) (Figure 1 B). The corrected p-values of the Tukey multiple mean comparison tests after ANOVA indicated that the patients with the GG genotype have a higher mean age of disease onset than the patients with the AA genotype (µGG – µAA = 7.414, padj= 0.046).
Bayesian logistic regression: the relationship of the OXTR gene variants with SASI and ASA
We examined the associations between OXTR rs237902, OXTR rs53576, and OXTR rs2254298 and the childhood and adulthood separation anxieties with respect to each demographic and clinical variable in the data, separately. In each Bayesian analysis, we first obtained posterior samples and then odds ratios and 90% credible intervals.
By examining the investigated genotypes in terms of the probability of separation anxiety using the SASI and ASA inventory results, we observed significant differences between the patients with the AA genotype and those with the AG or GG genotype. Strikingly, the patients with the AA genotype were found to be more likely to have childhood and adult separation anxiety than other genotypes in the presence of several entities (Tables II and III).
Table II
Relationships between OXTR gene variants and SASI scores
Table III
Relationships between OXTR gene variants and ASA scores
OXTR gene variants and SASI
When considering medium or high socioeconomic status and the OXTR rs237902 gene variant together, the patients with the AA genotype are OR = 1/0.335 = 2.99 times more likely to have childhood separation anxiety than patients with the AG genotype. For this gene variant, we observed that patients with the nocturnal panic and the AA genotype are OR = 1/0.159 = 6.29 times more likely to have childhood separation anxiety than patients in the same group with the AG genotype (Table II, column 2).
For OXTR rs53576 and married patients, the probability of having childhood separation anxiety is OR = 4.97 times higher for the GG genotype than the AA genotype. For OXTR rs53576, we observed that single patients with the AA genotype are OR = 1/0.033 = 30.30 and OR = 1/0.027 = 37.04 times more likely to experience childhood separation anxiety than single patients with the AG and GG genotypes, respectively. For the same gene variant, we observed that the probability of having childhood separation anxiety in unemployed patients with the AA genotype is OR = 1/0.046 = 21.74 and OR = 1/0.040 = 25 times higher than those with the AG and GG genotypes, respectively. In the patients without an anxiety-triggering life event, this probability is OR = 1/0.042 = 23.81 and OR = 1/0.034 = 29.41 times higher for those with the AA genotype than those with the AG and GG genotypes, respectively (Table II, column 3).
For OXTR rs2254298, in the case of a family history of psychiatric disease caused by panic disorder, the probability of childhood separation anxiety for the patients with the AA genotype is OR = 1/0.010 = 100 times more likely than those with the AG genotype (Table II, column 4).
OXTR gene variants and ASA
By examining OXTR rs237902 in terms of the probability of adulthood separation anxiety using the ASA inventory results, we observed significant differences between the patients with the AA genotype and those with the AG or GG genotype. In some cases, the patients with the AA genotype were found to be more likely to have adult separation anxiety than those with the AG genotype. The corresponding odds ratios were as follows: OR = 1/0.027 = 37.04 in female patients, OR = 1/0.026 = 38.46 in unemployed patients, OR = 1/0.257 = 3.89 in patients with medium or high socioeconomic status, OR = 1/0.027 = 37.04 in patients with nocturnal panic, OR = 1/0.190 = 5.26 in patients with an anxiety-triggering life event, and OR = 1/0.026 = 38.46 in patients with object carrying behavior. Similarly, the odds ratios indicating that the patients with the AA genotype are more likely to have adulthood separation anxiety than those with the GG genotype are as follows: OR = 1/0.046 = 21.74 in female patients, OR = 1/0.022 = 45.45 in unemployed patients, OR = 1/0.040 = 25 in patients with nocturnal panic, and OR = 1/0.060 = 16.67 in patients with object carrying behavior (Table III, column 2).
For OXTR rs53576, the probability of having adulthood separation anxiety was found to be higher in some cases for the patients with the AA genotype than those with the AG genotype. The significant odds ratios are as follows: OR =1/0.055 = 18.18 in single patients, OR = 1/0.032 = 31.25 in patients with nocturnal panic, and OR = 1/0.052 = 19.23 in patients without an anxiety-triggering life event. Similarly, this probability was found to be higher in some groups for the patients with the AA genotype than those with the GG genotype. The significant odds ratios are as follows: OR = 1/0.042 = 23.81 in single patients, OR = 1/0.043 = 23.26 in unemployed patients, OR = 1/0.058 = 17.24 in patients with nocturnal panic, and OR = 1/0.057 = 17.54 in patients without an anxiety-triggering life event (Table III, column 3).
For OXTR rs2254298 and the patients without an anxiety-triggering life event, the probability of having adulthood separation anxiety is OR = 1/0.038 = 26.32 times higher when comparing the AA genotype against the AG genotype. For the same gene variant and the patients with a family history of psychiatric disease caused by panic disorder, the probability of having adulthood separation anxiety is OR = 1/0.030 = 33.33 times higher for the AA genotype compared to the GG genotype (Table III, column 4).
Discussion
OXTR variations have been associated with behavioral patterns and predisposition to psychiatric diseases. To the best of our knowledge, there has been only one study which evaluated the effects of OXTR variants in panic disorder. In this study, the possible relationships between two OXTR variants (rs2254298 and rs53576) and panic disorder, social anxiety disorder, and major depressive disorder were investigated in Japanese participants. The G allele of OXTR could render the people more susceptible to panic and major depressive disorder [29]. Though we did not find a statistically significant association for rs2254298 and rs53576, OXTR rs237902 analysis revealed a significant association partially contrary to Onodera et al. [29]. Presence of the A allele was associated with a 1.585-fold increased probability for panic disorder.
Most of the studies have focused on SASI, while only a few have covered ASA. We evaluated the possible associations of the genotypes with SASI and ASA in a combined approach model covering several demographic and clinical data (Tables II and III). Genetic liability to complex traits could be better analyzed in the context of environmental and social factors, which are markedly linked to genetic liability. All of the analyzed variants displayed significant associations with several traits. The combinations of AA genotype of the analyzed variants + several other risk factors (Tables II and III) were associated with SASI and ASA scores. Although the studied models are not the same, our results seem partly discordant with the studies of Costa et al. [17], Schiele et al. [30], and Costa et al. [16] since the GG genotype effect was more prominent in these studies. On the other hand, the potential detrimental effect of the rs2254298 A allele in the presence of an environmental factor, as in the case of childhood emotional neglect [31] or childhood maltreatment [20], seems to be supported by our results. Nocturnal panic attacks which increase the severity of the disease were reported in half of the panic disorder patients [32]. Compatible with the more detrimental potential of AA genotype, OXTR rs237902 AA genotype in panic disorder patients with nocturnal panic attacks was found to be associated with SASI scores (Table II). Both OXTR rs237902 AA and OXTR rs53576 AA genotypes in panic disorder patients with nocturnal panic attacks were found to be associated with ASA scores (Table III). It seems likely that AA genotypes of the OXTR gene in the presence of nocturnal panic attacks worsen the course of the disease. The only exception referring to the effect of the GG genotype in our study was the combined analysis of OXTR rs53576 with marital status (Table II). While the probability of childhood separation anxiety for married people with the GG genotype was 4.97-fold compared with the AA genotype group, single patients with the AA genotype had 30.30-fold and 37.07-fold increased risks compared with the AG and GG genotypes, respectively. The GG genotype of married people seems to reflect a lower likelihood of childhood separation anxiety. This finding indirectly seems to support the results of Monin et al. [8], who drew attention to the association between GG genotype (self or partner effect) and marital satisfaction.
In conclusion, our study has many powerful dimensions, although our study design also had a notable limitation, as it involved the analyses of three OXTR variants; enriched studies with much more variants would be desirable. Despite this limitation, our study analyzed the effects of OXTR variants and developed one of the most comprehensive models in panic disorder research in terms of gene-environment (G × E) interactions. The enrichments of our findings would better enlighten the genetic underpinnings of PD and provide early intervention implications for clinicians to identify at-risk clinical subgroups. Since defects of the oxytocin system are reported in various psychiatric disease models, intranasal oxytocin use for therapeutic purposes might be on the agenda as an innovative approach in future.