Introduction
Ischaemic cardiovascular disease is a leading cause of mortality worldwide and is a major mortality threat in North America and South Korea [1, 2]. Of the various factors proposed to be responsible for the development of cardiac diseases leading to heart failure, ischaemia is the dominant one [3]. Ischaemia is characterised by hypoxia, apoptosis, and cell necrosis, which finally leads to organ dysfunction manifested as cardiac failure [4]. The ischaemic conditions cause a reduction in the blood flow due to arterial occlusion, which is responsible for the induction of endoplasmic reticulum (ER) stress [5]. ER stress produces reperfusion in the tissues affected and leads to production of reactive oxygen species (ROS) [6]. The ROS production coupled with reduced levels of intracellular antioxidants affect various cellular signalling pathways activating the apoptosis of cardiomyocytes, which may ultimately be presented as cardiac failure [7]. Hence, any agent with antioxidant potential, and which thereby can decrease the ROS levels and regulate the ER stress, may prove helpful in preventing ischaemia/reperfusion-induced heart failure. Natural compounds act as a reservoir for the discovery of such compounds. The natural compounds as depicted by recent research show a number of beneficial effects to human health [8]. They have been shown to possess tremendous antioxidant potential and bear anti-apoptotic effects [9]. The same properties are being exhibited by the flavonoid class of natural compounds [10]. The flavonoid compounds have attained huge scientific attention due to their vast pharmacological properties [11]. These compounds function as anti-inflammatory, anti-microbial, and anti-carcinogenic agents [12]. Previous studies have shown that flavonoid-based compounds are active in alleviating oxidative stress and have cardiac preventive roles [13]. Considering this, the present study was devised to unravel the effects of tangeretin, a flavanone compound, in preventing the oxidative stress mediated by ischaemia/reperfusion, and thereby to analyse its cardiac protective effects in terms of its anti-apoptotic potential on the heart and the underlying molecular mechanisms thereof. The results showed that tangeretin inhibited the apoptosis of cardiomyocytes and enhanced their viability by modulating the Bax/Bcl-2 protein ratio.
Further, the anti-apoptotic property of tangeretin was further manifested as the decline of caspase-3 activity. Taken together, tangeretin may have therapeutic implications in the treatment of cardiac diseases.
Material and methods
Ischaemia/reperfusion induction in cardiomyocytes
To perform the stimulation of ischaemia/reperfusion in vitro, the rat H9c2 cardiomyocytes were brought from Vital River Laboratories, China and maintained in DMEM medium at 37°C in an anaerobic glove box (Coy Laboratory, USA) from which normal air was removed by a vacuum pump and replaced with 5% CO2, 5% H2, and 90% N2. The H9c2 cardiomyocytes were cultured at a density of 5 × 104 cells/ml under hypoxia for 4 h. Then, the cells were removed from the anaerobic glove box, and the medium was replaced with high glucose medium and maintained in the regular incubator to mimic reperfusion.
Analysis of cell viability
The normal cardiomyocytes, untreated and tangeretin-treated ischaemic H9c2 cell lines, were induced were cultured to a density of 5 × 103 cells/well in 96-well plates and treated with 9 µM concentration of tangeretin (in 0.1% DMSO) for different time periods (0, 12, 24, 48, and 96 h). The assessment of cell viability was made by using the Cell Counting kit-8 (CCK8), following the manufacturer’s guidelines.
RNA isolation, cDNA synthesis, and qRT-PCR
Total RNA from the normal cardiomyocytes, untreated, and tangeretin-treated ischemic H9c2 cell lines was extracted by Trizol method. Exactly 2 µg RNA was reverse-transcribed to cDNA with a SuperScript First-Strand Synthesis System (Thermo Scientific). The qPCR analysis was performed to determine the expression levels of the genes of interest through the SYBR Green method. The expression levels were quantified using the 2–ddCt method. To normalise the expression, human GADPH was used to serve as an internal expression control.
Assessment of antioxidant potential of tangeretin
The antioxidant potential of tangeretin was investigated by harvesting rat ischaemic cardiomyocytes administered with 0, 4.5, 9.0, and 18 µM concentrations of tangeretin by estimating the intracellular levels of superoxide dismutase (SOD), catalase (CAT) enzymes, and reduced glutathione (GSH). The estimations were made using the respective detection kits.
5-ethynyl-2′-deoxyuridine assay
A 5-ethynyl-2′-deoxyuridine (EdU) Kit (C00031Apollo®567 was used to determine the proliferation rate of the cardiomyocytes. Briefly, the H9c2 cells were subjected to culturing at 25°C. This was followed by the addition of EdU solution to the cell culture for 2 h. The H9c2 cells were then subjected to fixation with paraformaldehyde (40 g/l) for 30 min. The H9c2 cells were then treated with glycine for 10 min and washed with phosphate-buffered saline (PBS). Next, the H9c2 cells were administrated with Apollo® staining solution at 25°C for 20 min without light. The H9c2 cells were also stained with 4′,6-diamidino-2-phenylindole (DAPI) at room temperature, and finally fluorescence microscopic analysis was performed to examine the cells.
Reactive oxygen species determination
The accretion of reactive oxygen species (ROS) in the rat cardiomyocytes was estimated with the help of 2′,7’-dichlorofluorescein diacetate (DCFH-DA). Briefly, the cell cultures were centrifuged at 25,000 rpm for 7 min and PBS washed. The H9c2 cells were again suspended in fresh media and treated with 10 mM DCFH-DA at 37°C incubation for 25 min. These H9c2 cells were then rinsed thrice in PBS, and the ROS levels were estimated by flow cytometry.
Western blotting
The quantitative assessment of the concentration of proteins of interest from the H9c2 cells, treated with 0, 4.5, 9.0, and 18 µM tangeretin, was made through western blot analysis. In brief, the cells were harvested and fractioned using the cell lysis buffer, and total protein concentrations of cell lysates were estimated through the Bradford method. Subsequently, the lysates with equal protein counts were loaded on 10% PAGE gel, which was then blotted to a PVDF membrane. The membrane was subsequently exposed to primary and secondary antibodies, and finally the chemiluminescence-based estimation of respective protein concentrations was made. Human GADPH protein was used as an internal control in the blotting experiments.
Results
Tangeretin enhanced the viability of ischaemic H9c2 cardiomyocytes
Tangeretin is a flavanone compound (Figure 1 A), whose molecular structure is presented in Figure 1. In order to analyse the effects of tangeretin in enhancing the cardiomyocyte viability, tangeretin (9 µM) was administered on ischaemic H9c2 rat heart cells. It was found that the tangeretin administration successfully enhanced the proliferation of H9c2 ischaemic cardiomyocytes to a considerable level, which was comparable to the normal cardiomyocytes (Figure 1 B). Nonetheless, the untreated ischaemic cardiomyocytes showed a very low proliferation rate.
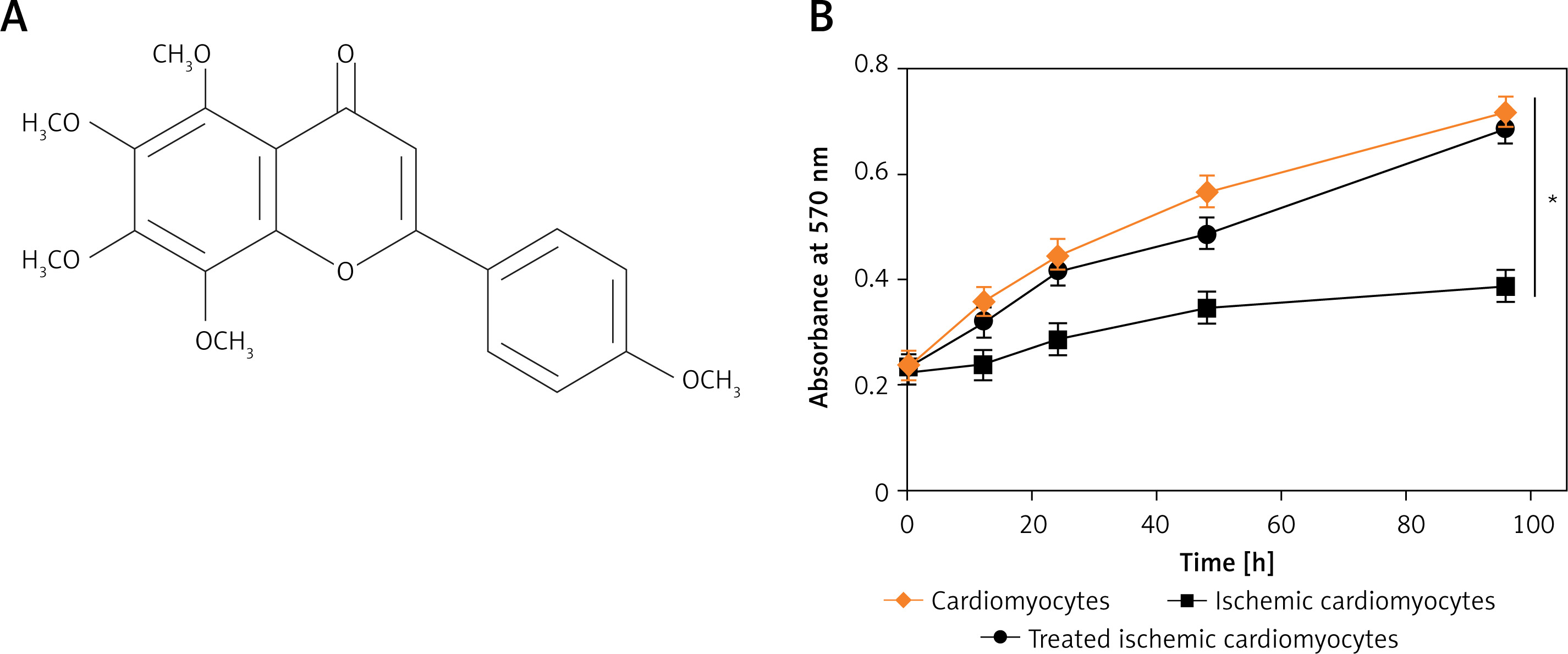
Tangeretin enhanced the antioxidant power of cardiomyocytes
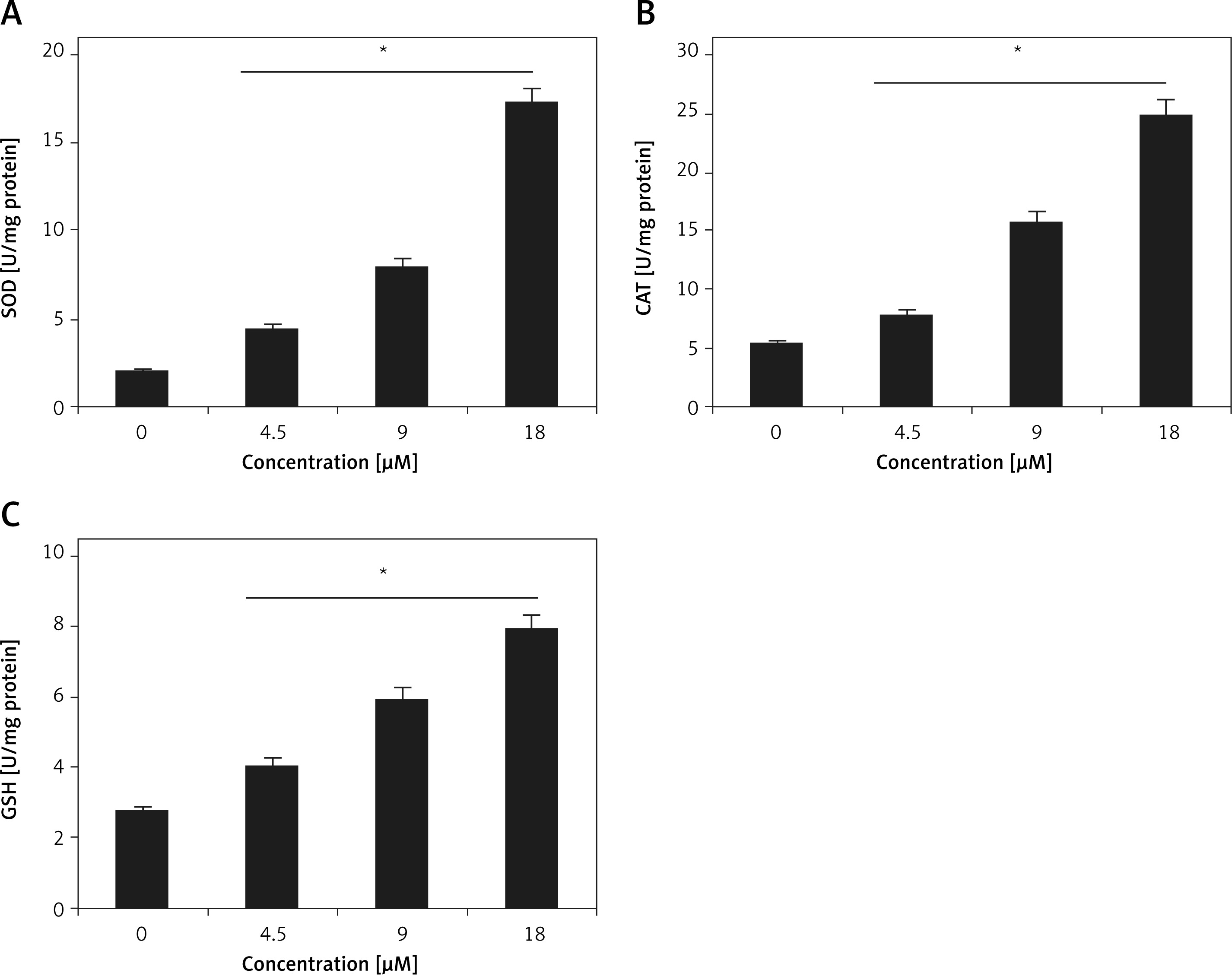
One of the important objectives of current study was to evaluate the antioxidant potential of tangeretin, and for elucidation of the same, the enzymatic (SOD and CAT) and non-enzymatic (GSH) anti-oxidant levels were estimated in cardiomyocyte cellular samples treated with 0, 4.5, 9.0, 18 µM tangeretin. It was found that the antioxidant levels in tangeretin-treated samples were significantly higher than those of untreated ones (Figures 2 A–C). The increase in antioxidant levels followed a concentration-dependent trend.
Tangeretin reduced the reduced apoptosis of H9c2 cells
The EDU and DAPI staining-based analysis of H9c2 cardiomyocyte, ischaemic cardiomyocytes, and tangeretin-treated ischaemic cardiomyocytes showed that the proliferation rates were decreased in ischaemic cardiomyocytes due to the induction of apoptosis. However, tangeretin treatment prevented apoptosis in the ischaemic cardiomyocytes because the proliferation rate of normal cardiomyocytes was comparable to that of the tangeretin-treated ischaemic cardiomyocytes (Figure 3 A).
Figure 3
A – EDU and DAPI staining showing proliferation in normal H9c2 cardiomyocytes, ischaemic cardiomyocytes, and tangeretin-treated ischaemic cardiomyocytes. B – Annexin V/PI assay showing apoptosis in normal H9c2 cardiomyocytes, ischaemic cardiomyocytes, and tangeretin-treated ischaemic cardiomyocytes. C – Western blot analysis showing Bax, Bcl-2, and caspase-3 expression in normal H9c2 cardiomyocytes, ischaemic cardiomyocytes, and tangeretin-treated ischaemic cardiomyocytes. The experiments were performed in triplicate and expressed as mean ± SD (*p < 0.05)
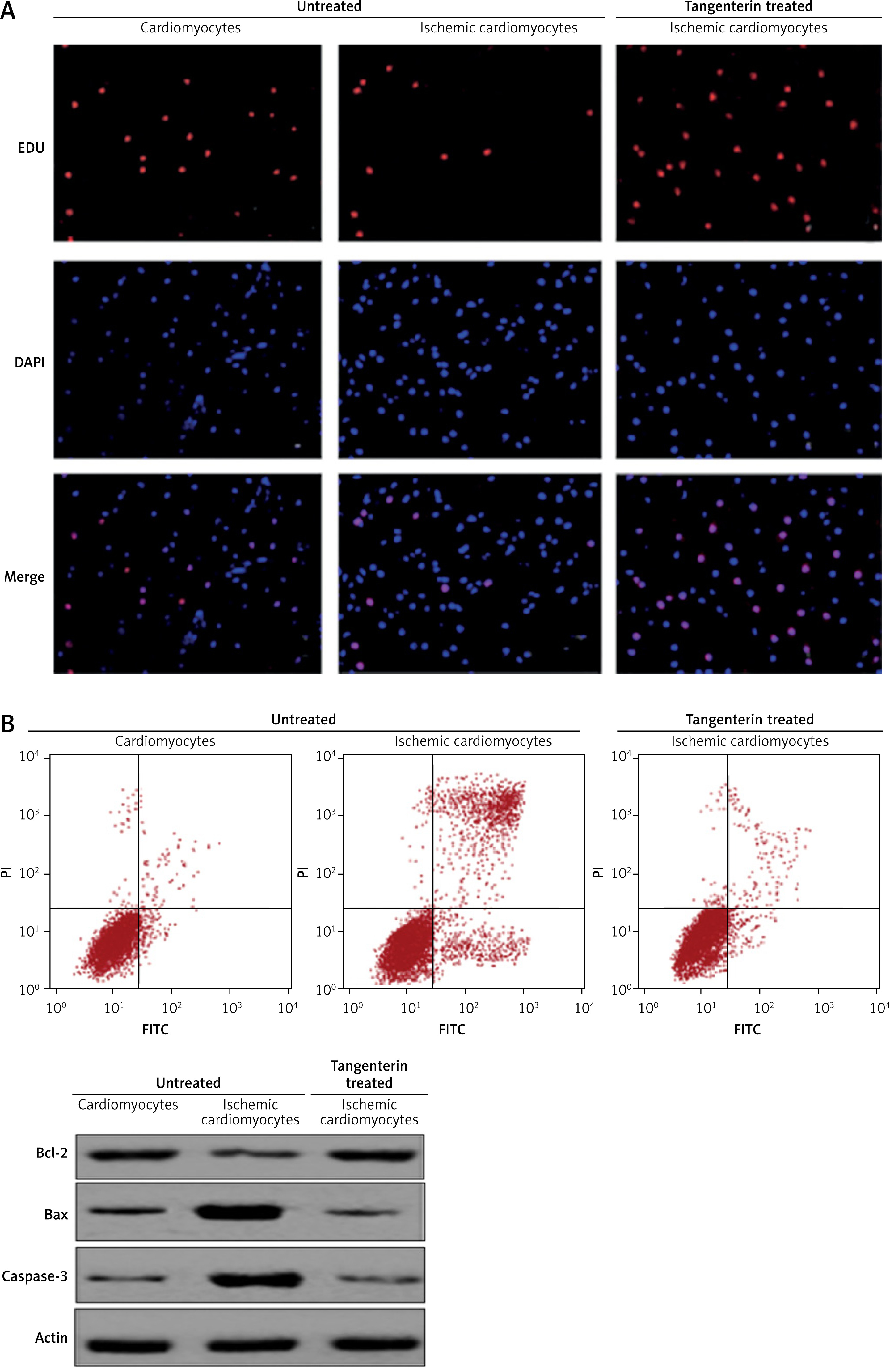
The annexin V/IP staining analysis showed that apoptosis was higher in ischaemic cardiomyocytes, while the apoptotic cell percentage was negligible in normal cardiomyocytes and tangeretin-treated cardiomyocytes (Figure 3 B). The results were further confirmed by the expression analysis of caspase-3, Bax, and Bcl-2 proteins. The expression of Bax and caspase-3 increased and Bcl-2 decreased in ischaemic cardiomyocytes. However, tangeretin treatment decreased the expression of Bax and caspase-3 and increased the expression of Bcl-2, thereby exerting protective effects by inhibiting apoptosis (Figure 3 C).
Tangeretin actively modulated the ER stress in ischaemic cardiomyocytes
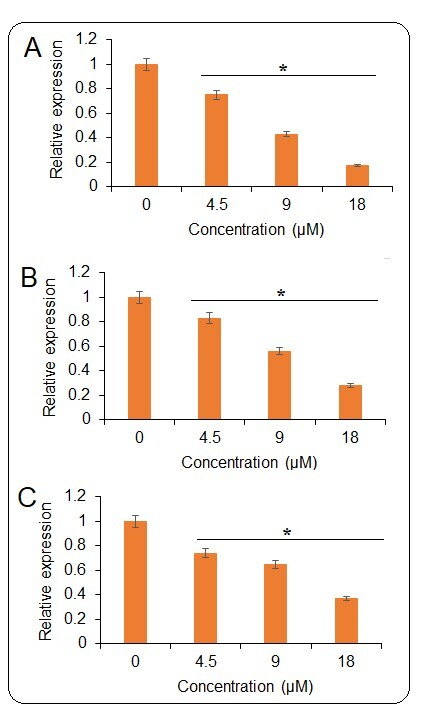
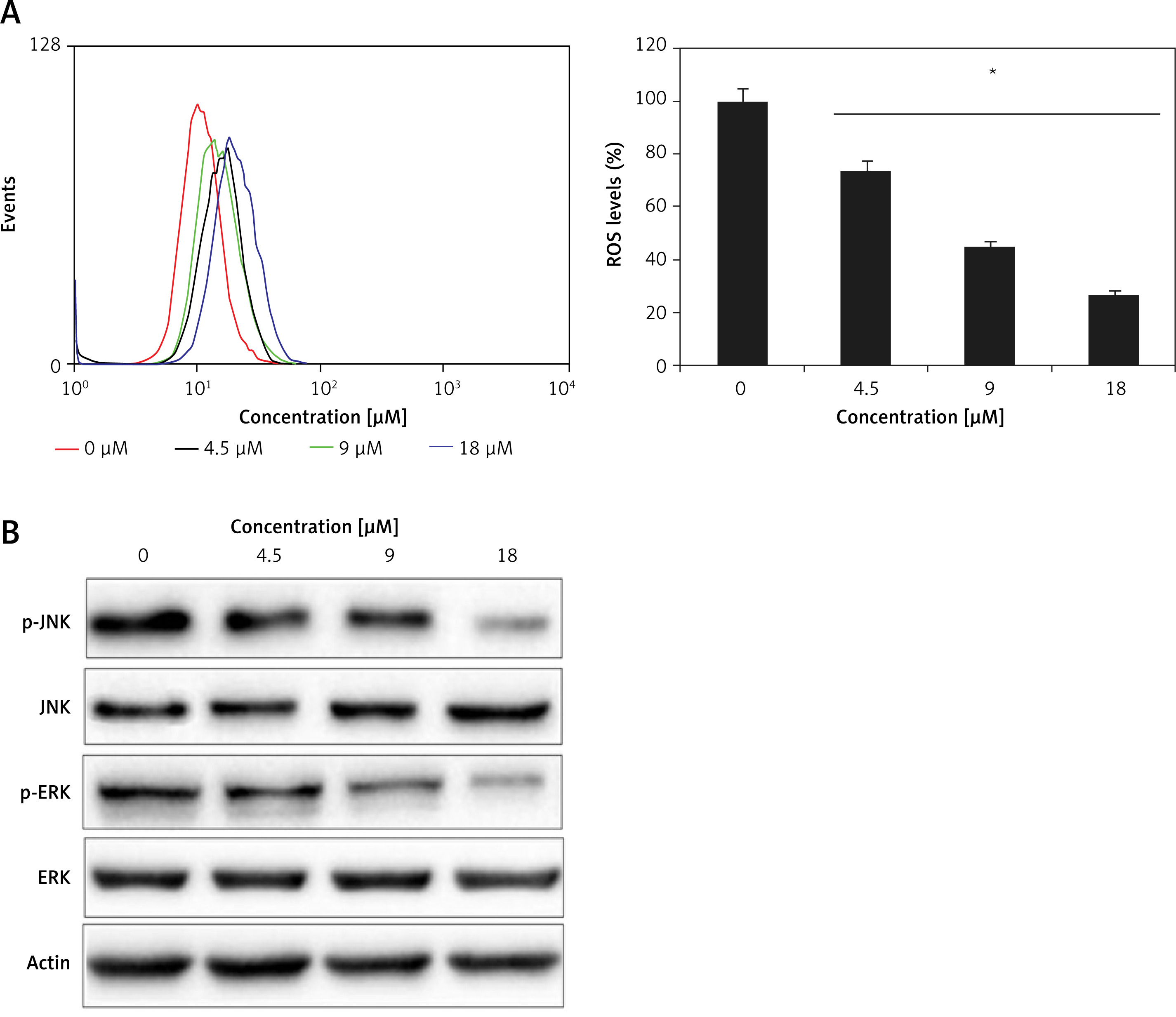
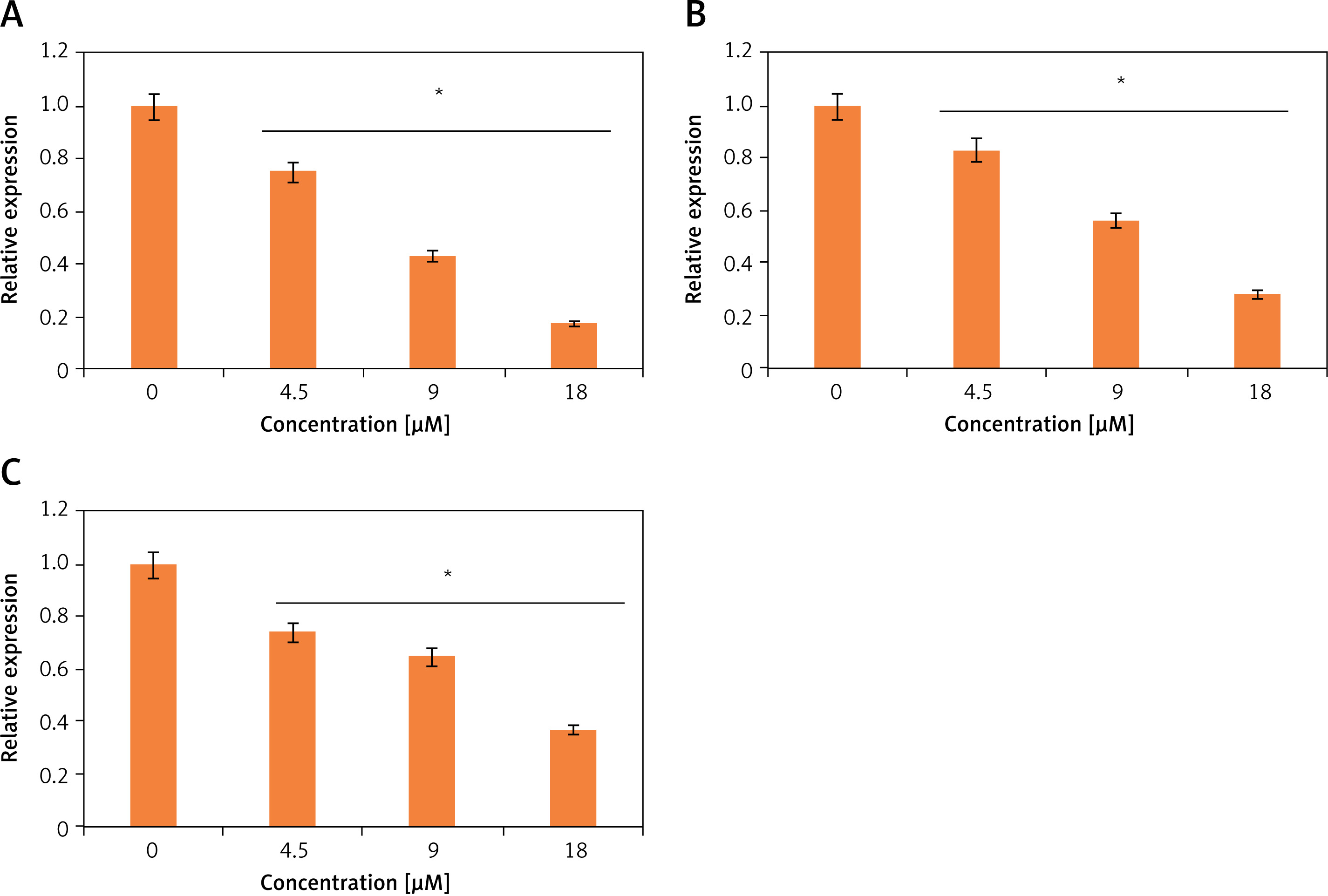
Ischaemic cardiomyocytes showed very high levels of ROS, and tangeretin treatment could effectively decrease the reactive oxygen species dose dependently (Figure 4 A). Ischaemia/reperfusion has been indicated to cause ER stress wherein different signalling pathways central to ER mediate the apoptosis of heart cells and ultimately result in heart failure. The JNK/ERK pathway is important in this regard. The ischaemic H9c2 cells exhibited higher levels of phosphorylated versions of JNK and ERK (p-JNK and p-ERK) proteins, which activate the transcription of downstream genes and lead to cell apoptosis. Interestingly, the tangeretin treatment decreased the p-JNK and p-ERK proteins (Figure 4 B). The protein level of JNK and ERK remained constant upon tangeretin treatment. Additionally, the expression analysis by qRT-PCR showed that the expression levels of the downstream genes such as c-fos, c-Jun, and c-myc decrease significantly and concentration dependently upon tangeretin treatment (Figures 5 A–C). Together these results indicate that tangeretin is active in modulating ER stress to enhance cell viability.
Figure 4
A – Flow cytometric analysis showing ROS levels in H9c2 ischaemic cardiomyocytes treated with different concentrations of tangeretin. B – Western blot analysis showing the effects of tangeretin on the JNK/ERK signalling pathway in H9c2 ischaemic cardiomyocytes. The experiments were performed in triplicate and expressed as mean ± SD (*p < 0.05)
Discussion
Ischaemia, which is characterised by the build-up of anoxygenic/hypoxic conditions in the human body, leads to the accretion of oxidising agents or ROS [5, 6]. ROS-mediated oxidative stress is a potent threat to cell viability [14]. Human cardiac cells have been shown to be comparatively susceptible to ischaemia-driven oxidative stress. Oxidative stress has lethal particularly effects on intracellular organelles like mitochondria and endoplasmic reticulum [15, 16]. In such a state, the normal organelle balance is disturbed, which leads to the activation of signalling cascades favouring the induction of apoptotic cell death. Herein, we examined the cardioprotective role of tangeretin, a flavanone on human cardiomyocytes. The results suggested that tangeretin-treated rat cardiomyocytes exhibited significantly higher cell viability. Such effects have been reported for other natural compounds in previous studies [17]. Tangeretin was seen to inhibit cardiomyocyte apoptosis markedly by inhibiting the activation of caspase-3. Caspase-3 activation leads to the activation of downstream apoptotic signalling components resulting in cell death [18]. Furthermore, the Bax/Bcl-2 ratio decreased considerably in the cardiomyocytes administered with tangeretin. Lower Bax/Bcl-2 is inhibitory to apoptotic cell death [19]. All these findings point towards the cardioprotective role of tangeretin which can be attributed to its anti-apoptotic effects. The findings of our study are also in agreement with a previous study wherein quercetin, a naturally occurring flavonoid, was shown to trigger cardioprotective effects by preventing the apoptosis of cardiomyocytes [20]. Increased ROS levels have been shown to activate apoptotic cell death in different cell types [21]. Herein, we observed that tangeretin significantly inhibited the generation of ROS and therefore prevented the apoptosis of cardiomyocytes. Furthermore, the tangeretin treatment was found to relieve ER stress by targeting the JNK/ERK pathway and by suppressing the expression of downstream genes. The phosphorylation of signalling components of this pathway are prime to triggering the suppression of downstream target genes, which results in the programmed cell death [22]. Thus, by blocking the phosphorylation of JNK and ERK proteins, the tangeretin actively diminishes the apoptotic cell signal. Previous studies have shown that production of ROS may lead to the activation of the JNK and ERK pathways [23]. For instance, cypermethrin has been shown to activate the JNK and ERK pathways in macrophages via induction of ROS production and consequent oxidative stress [24]. Thus, inhibition of the JNK and ERK may be attributed to the suppression of ROS generation by tangeretin. Nonetheless, we do not overrule the possibility of direct inhibition of JNK and ERK pathway by tangeretin. Altogether, the results of this study highlight the cardioprotective potential of the natural compound tangeretin and advocate further confirmation by performing evaluation of tangeretin on more cell lines and under in vivo conditions in animal models.
In conclusion, the present study provides insights about the protective role of tangeretin on cardiomyocytes via suppression of ROS, inhibition of apoptosis, and deactivation of the JNK and ERK pathways. Although these findings point towards the cardioprotective role of tangeretin, in vivo studies in animals and in humans are required for further confirmation.