Alzheimer’s disease (AD) is an irreversible and progressive neurodegenerative disorder characterized by the deposition of β-amyloid, intra-neuronal presence of neurofibrillary tangles, glial hypertrophy, and intracellular accumulation of hyperphosphorylated tau implicated in memory and learning deterioration. It has been hypothesized that dysregulation of Ca2+ disrupts the synaptic network in AD [1–3]. Synapse loss is associated with increased endoplasmic reticulum (ER) Ca2+ release and Ca2+ hyperactivity [4]. Cytosolic Ca2+ levels are tightly regulated and maintained at low concentrations, despite a much higher extracellular Ca2+ concentration and similar regulation to an elevated Ca2+ level within the ER. Ryanodine receptor 3 (RYR3) have a role in gating ER Ca2+ and are sensitive to changes in cytosolic Ca2+. The changing cytosolic Ca2+ levels activated the RYR channel and inositol 1,4,5-triphosphate receptor (IP3R) as one of the synaptic processes to maintain Ca2+ homeostasis in multiple neuronal functions [3, 4]. In presenilin-1 (PS-1)-expressing AD transgenic mouse increased RYR expression was observed followed by enhancement of ryanodine-Ca2+ release. The same results also showed a caffeine-induced ryanodine-mediated Ca2+ response, which is aggravated in triple-transgenic mouse models (3xTg-AD) [5–8].
Aluminum (Al) is a neurotoxin that leads to development of symptoms similar to those for AD such as depression and cognitive deterioration. Aluminum increased enzyme activities such as catalase, superoxide dismutase (SOD), and glutathione reductase (GR). In vitro studies showed that Al can promote Aβ aggregation and neurotoxicity by induction of glial activation and initiation of macrophage responsivity, promoting inflammation of glial cells. Subsequent studies reported systemic aluminum-induced AD-like behavioral deficits in treated rats that led to increased levels of Aβ1-42 [9, 10].
Many studies have shown that the promising findings obtained from preclinical studies provide strong results presenting mesenchymal stem cells (MSCs) as a potential alternative therapy in humans. MSCs are described as non-hematopoietic, multi-potent stem cells isolated from umbilical cord blood, Wharton’s jelly of the umbilical cord, placenta, amniotic fluid, and adipose tissue. Transplanted MSCs release several trophic factors into the host environment, reducing inflammation, apoptosis, and fibrosis of damaged tissues [8]. Studies have shown that MSCs stimulate neurogenesis in neurodegenerative diseases. An in vitro study using primary mesencephalic culture with protease inhibitor treatment with MSCs showed increased dopaminergic neuronal survival and a significant reduction of caspase-3 activity.
Moreover, MSCs can interact with the resident cells in the brain and further regulate the homeostatic response to promote tissue preservation and functional recovery by producing neurotrophins, vascular endothelial growth factor, nerve growth factor, and anti-inflammatory cytokines [11]. In AD, MSC therapy showed attenuation of β-amyloid deposition, up-regulation of interleukin-10 (IL-10), and down-regulation of interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α). A previous study reported that MSCs co-cultured with Chinese hamster ovary (CHO) cells stably expressing human amyloid precursor protein (APP) 695 showed markedly enhanced autolysosome formation and clearance of Aβ in AD models, leading to increase neuronal survival against Aβ toxicity, repair of the damaged AD brain and increased survival of hippocampal neurons [11, 12].
RYR genes are known as intracellular calcium release channels and regulate the function of synapse plasticity, neuronal viability, and cognitive functions. RYR3 is expressed in the granular cell layer of the cerebellum, hippocampus, thalamic, hypothalamic, caudate, and putamen nuclei. Like other RYR families, RYR3 has been reported for its role in the calcium signaling pathway. A previous study showed that RYR3 expression increased in the hippocampus of APP/PS1 mice at age 3 months and 6 months [7, 8, 13]. This gene stimulates Ca2+ release independently from inositol 1,4,5-triphosphate (IP3), increasing cytosolic calcium concentration [7, 8, 14].
Thus, the current study aims to observe the effect of MSCs in attenuating β-amyloid deposition and its association with the RYR3 gene in AD rats.
Methods
Ethical approval
This study was conducted after approval by the Ethics Committee of the Faculty of Medicine, Andalas University, Indonesia, with approval number 796/UN.16.2/KEP-FK/2022.
Culture of mesenchymal stem cells (MSCs)
Wharton’s jelly derived mesenchymal stem cells were obtained from Stem Cell and Tissue Engineering (SCTE), Indonesian Medical and Research Institute (IMERI), Faculty of Medicine, University of Indonesia. MSCs were plated at 100 × 104 cells/cm2 and expanded in vitro in α-Modified Eagle’s medium (α-MEM; Gibco, USA) containing 10% fetal bovine serum (FBS; Sigma Aldrich, USA) and 1% penicillin/streptomycin (Gibco, USA) in a standard 5% CO2 incubator at 37 ºC. Cells from P5 and P6 were used in all experiments.
Animals
Twenty male adult Wistar rats (6–8 weeks of age) weighing 200 ±20 g were obtained from the university Animal House and were distributed randomly into three groups of animals (n = 5). The experimental procedures were carried out in accordance with the international guidelines for the care and use of animals. Aluminum chloride (AlCl3) purchased from Millipore Merck, USA, cat.no.1.01084.1000, was used to induce neurotoxicity in AD as previously described [15]. The AlCl3 group was given 300 mg of AlCl3/kg body weight (BW) intraperitoneally (i.p.) for 5 days. The combination AlCl3 and MSC group received an MSC injection and was observed after 30 days.
Congo red stain for the amyloid β and Y-maze test
Sections were deparaffinized and rehydrated as previously described, and a Y-maze test was carried out to observe the mice’s willingness to discover the new environment.
Realtime PCR
cDNA was synthesized from 1 µg of RNA using a SensiFAST cDNA synthesis kit (Bioline, USA). Real-time PCR amplification reaction using the SensiFAST real-time PCR Kit (Bioline, USA) was performed using the Bio-Rad CFX96 Real-time PCR Detection System (Bio-Rad, USA) to detect RYR3. Primers for RYR3 detection were 5′-TGGGACAAGTTCGTGAAG-3′ for forward and 3′-GCCCATGGCAATATCCAA-5′ for reverse. The housekeeping gene of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control with forward 5′-AGAAGGTGGTGAAGCAGGC-3′, reverse 3′-GTCCACCACCCTGTTGCTG-5′.
Statistical analysis
Data are presented as means ± SEM. Results of Congo red staining and RYR3 expression were analyzed using the one-way ANOVA test followed by the Tukey-Kramer test for post hoc analysis to examine further the significant differences in the means between the groups. P-values < 0.05 were considered statistically significant. All statistical analyses were performed using the SPSS v.22 software (SPSS Inc., Chicago, USA).
Results
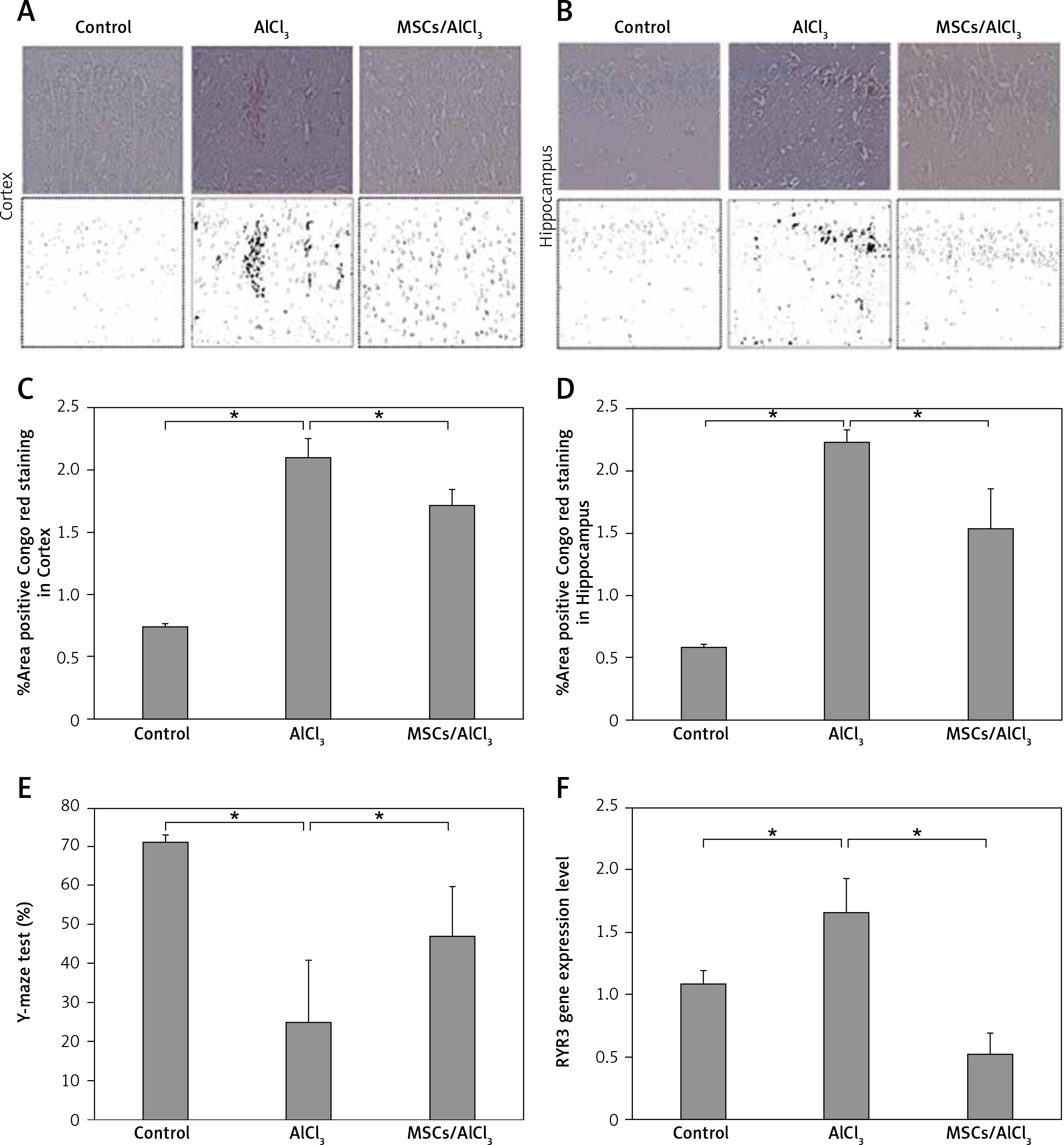
The injection of AlCl3 induces amyloid deposition in the cortex and hippocampus of this group. Congo red staining was detected in AlCl3-induced mice, and lower amyloid accumulation was found in AlCl3-MSCs treated mice 30 days after MSC injection (Figures 1A–D).
Figure 1
Amyloid deposition in the cortex and hippocampus was detected in AlCl3-induced AD rats. MSCs improved the amyloid deposition 30 days after treatment (A–D). Y-maze scores were enhanced 30 days after MSC treatment (E). Interestingly, MSC treatment exhibited lower expression of the RYR3 gene, suggesting improvement in Ca2+ regulation (1F); *p < 0.05
RYRs are known for their role in Ca2+ regulation. The current results showed that the expression of RYR3 was noticeably increased after the injection of AlCl3. Interestingly, 30 days after MSC treatment, the presentation decreased. This result is similar to the previous study, which showed that lower RYR3 expression improved the AD condition.
Discussion
The current study showed the deterioration of the cortex and hippocampus region in the brain after injection of AlCl3 intraperitoneal. The duration and 300 mg/kg BW of AlCl3 administration induced AD symptoms based on the previous findings according to the neurotoxicity dose [9, 10]. Amyloid deposition is higher in AD rats’ cortex and hippocampus than in other groups. AlCl3 induction appears to cause selective changes in transport systems in the blood-brain barrier (BBB), further inducing oxidative stress by damaging the membrane lipid, inflammation, and disrupting neurotransmitters in the brain hippocampus, further altering physiological function and leading to brain disorder, inducing programmed cell death, vacuolar spaces, distortion in the histoarchitecture and loss of the Purkinje cell layer of the cerebellar cortex [15, 16].
The current study used MSCs derived from Wharton’s jelly of the umbilical cord. They have been reported for their function in the renewal of dying cells within the tissues by the differentiation of stem cells. MSCs are known for their strong immunomodulatory properties and strong paracrine properties and secrete various cytotropic factors that, in turn, exert neuroprotective effects. Our study showed that amyloid accumulation in the cortex and hippocampus was lower in the MSC-treated group, 1.72 ±0.13 and 1.54 ±0.31 (p < 0.05), respectively, compared to the AlCl3 group without MSCs. Previous animal studies demonstrated that human MSCs have neuroprotective effects through complex mechanisms, such as modulation of inflammation, enhancement of cell survival signals, increased neurogenesis, and modulation of ubiquitinated proteins [11, 12].
In AD, the perturbation of endoplasmic reticulum (ER) Ca2+ homeostasis has been linked to the cleavage of amyloid precursor protein (APP) by β-secretase and presenilin-1 (PS1), further generating amyloid β peptides. RYR3 presents highly in the brain. This gene has been reported to increase AD and mediated ER Ca2+ alteration. A previous study showed that RYR3 expression increased in PS1 knock-out mice, suggesting increased Ca2+ release through RYR in the pathogenic AD mechanism [7, 8, 17]. The present study showed that RYR3 expression is lower, 0.52 ±0.171 (p < 0.05), compared to AlCl3 mice without MSC treatment, 1.66 ±0.268 (p < 0.05) (Figure 1 F). MSC administration significantly reduced the amyloid deposition in the cortex and hippocampus, further lowering RYR3 expression. It is well documented that alterations in Ca2+ homeostasis occur early in AD before rigorous neuronal damage and cell death are noticed. The changes in Ca2+ levels were postulated to correlate more closely with cognitive decline compared with amyloid-β accumulation in senile plaques [17–19].
This study is the first to report that the MSC-treated AlCl3 group showed lower RYR3 expression. Our study strengthens the benefits of MSCs for AD treatment. Future research should examine whether MSCs improve AD and Ca2+ regulation in the clinical setting.
In conclusion, MSCs improved amyloid accumulation in AD animal models. The lower amyloid accumulation also lowered the RYR3 expression. MSCs might play a role in Ca2+ homeostasis, further alleviating learning ability in AD animal models.