Introduction
Osteosarcoma (OS) is the most common primary bone tumor [1], and the main affected population is adolescents [2–4]. The survival of OS patients was 10–20% when surgery was used as a single treatment. After the implementation of comprehensive treatment in 1985, the survival rate increased to 50–60%, and since then the survival rate has not significantly improved [2, 5–10]. There is less basic research on OS than for other tumors, and we need more ways to improve the survival rate.
The human phosphotyrosine picked threonine kinase (TTK) gene, also knowns as monopolar spindle 1-like 1 (MPS1), is located on chromosome 6q13-q21. The kinase family of TTK is 85–95 kDa and has conserved C-terminal domains [11]. TTK/MPS1 encodes an important protein kinase, which is essential in spindle checkpoint [12, 13], chromosome segregation [14] and chromosome stability and localized to kinetochores [15]. In addition to its mitosis function, TTK is involved in DNA damage repair through phosphorylation of p53, CHK2, and MDM2 [16]. For decades, TTK has been widely reported as an oncogene in multiple types of cancers, such as hepatocellular carcinoma (HCC) [17–19], and triple-negative breast cancer [20]. Phosphotyrosine picked threonine kinase is enhanced in triple-negative breast cancer compared to the other breast cancer subgroups at the protein level and the inhibition of TTK severely compromises viability of breast cancer cells and their capacity to form colonies in an anchorage-independent manner [21].
Phosphotyrosine picked threonine kinase is also known as a clinical therapeutic target. NTRC 0066-0, an inhibitor of TTK with subnanomolar IC50, shows an increase in the survival of a murine breast cancer model that mimics human triple-negative breast cancer (TNBC) and delays tumor relapse in combination with docetaxel [22]. Thus the combination treatment of TTK inhibitor and taxane chemotherapy has been well developed and entered the phase I dose escalation study [14]. Additionally, TTK is also involved in the oxidative damage repair of DNA and regulated cell survival [23]. Notably, TTK could contribute to the formation of blood vessels in tumors. The inhibition of TTK in tumors decreased vascular density and improved survival, which was confirmed by a lung cancer animal model [24]. Epithelial-to-mesenchymal transition (EMT) in TNBC cells could be reversed by pharmacologic inhibition and genomic silencing of TTK and the expression of KLF5 acting as an effector of TGF-β signaling and inhibitor of EMT could also be increased by this strategy [22].
This study aims to assess TTK expression levels in human OS tissues and its link with the clinical characteristics of OS patients, and to evaluate the potential role in OS development. In this study, we found abnormally high expression levels of TTK in human OS tissues and identified the potential link between TTK expression and the clinical pathological features of OS patients. We further found that TTK knockdown dramatically restrained OS cell proliferation and suppressed tumor growth of OS cells in mice. Collectively, TTK could therefore be considered as a possible therapeutic target for the treatment of OS.
Material and methods
Specimen collection
Based on the institutional review board’s approval, this retrospective study identified 74 consecutive patients with OS who underwent pancreatic cancer resection in our hospital between 2013 and 2018. Patients were classified according to the 2009 UICC TNM staging and the 2004 WHO/ISUP classification. All patients had no preoperative radiotherapy or chemotherapy. Attendees provided written informed consent. All patients showed no evidence of tumor metastasis confirmed by cross-sectional imaging.
Cell line culture
Osteosarcoma cell lines MG-63 and U-2 OS were purchased from ATCC (Manassas, VA, USA) and were maintained at 37°C with 5% CO2. According to the recommendations of ATCC, MG-63 cell lines were cultured with DMEM (11965-092, Gibco, Waltham, USA) with 10% FBS (A31605, Gibco, Waltham, USA), 1% Penicillin-Streptomycin (P1400, Solarbio, Beijing, China) and U-2 OS cell lines were cultured in McCoy’s 5a Medium Modified (16600082, Gibco, Waltham, USA) with 10% FBS, 1% Penicillin-Streptomycin.
Immunohistochemistry
The patient and animal tissues were fixed in formalin and embedded in paraffin, and 4 µm – thick sections were cut from the paraffin tissues. Sections were heated in a microwave for 20 min at 95°C in citrate buffer to retrieve the antigen, and they were washed three times with phosphate-buffered saline (PBS) buffer after cooling at room temperature. Subsequently, immunohistochemical (IHC) assay was performed according to the manufacturer’s instructions. Briefly, endogenous peroxidase was blocked using 3% (v/v) hydrogen peroxide for 30 min at room temperature, and the sections were washed three times in PBS, blocked using 2% bovine serum albumin for 1 h at room temperature, then incubated with primary antibodies overnight at 4°C in a humidified chamber. Finally, the sections were washed three times, incubated with second antibody-horseradish peroxidase for 1 h at 37°C and colors were developed with hematoxylin and 3,3-diaminobenzidin. Pictures were collected using a microscope.
Colony formation assay
The cells were digested with trypsin and the cell density was adjusted to 1000 cells/ml. 2 ml of complete medium with cells was added to each well of a 6-well plate, and cultured in a 37°C, 5% CO2 incubator for 2 weeks. After 2 weeks, the medium was aspirated, fixed with 4% paraformaldehyde for 15 min, washed three times with PBS, and then stained with crystal violet for 15 min, and they were photographed after being washed in crystal violet with PBS.
Cell proliferation assay
Cells were seeded into 96 wells with complete medium, and each well contained 5000 cells in 200 µl of medium. The 96-well plates were taken out every 24 h to evaluate the cell proliferation using the cell counting kit-8 (CCK8) (96992, Sigma, Saint Louis, USA) assay according to the manufacturer’s instructions. Simply, 20 µl of CCK8 was added into each well and cultured in a cell incubator for 4 h, and the absorbance was evaluated using a microplate reader at 450 nm wavelength (Bio-Rad, Hercules, USA).
Western blotting
Cells were washed three times with cold PBS and lysed in RIPA (89900, Thermo, Waltham, USA) containing 1% PMSF (36978, Thermo, Waltham, USA), and then the cells were incubated on a shaking table in 4°C for 30 min. The lysate was centrifuged at 12000 r/min for 30 min at 4°C. 30 µg of protein, the concentration determined using Bradford assay (P0006, Beyotime, Shanghai, China), was electrophoresed by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (P1200, Solarbio, Beijing, China) at 90 V, and then transferred to PVDF (ARKEMA, Colombes, France) membranes at 250 mA for 130 min. The membranes were blocked with 5% non-fat milk for 1 h at room temperature after being washed three times with tris-buffered saline and Tween (TBST). Then, the PVDF was incubated overnight with primary antibodies on a shaking table at 4°C. Primary antibodies were: TTK (ab219068, 1 : 1000, Abcam, Cambridge, UK), β-actin (1 : 5000, KM9001T, Sungene, Tianjin, China), ki67 and proliferating cell nuclear antigen (PCNA) (1 : 1000, 9449T, 2586S, Cell Signaling Technology, Danvers, UK). Then, the second antibodies (1 : 10000, LK2003, Sungene, Tianjin, China) were incubated for 1 h on a shaking table at 37°C after washing three times with TBST. Finally, ECL reagents (35050, Thermo, Waltham, USA) were added to display strips and take pictures using an image capture system.
Knockdown phosphotyrosine picked threonine kinase
Lentivirus was used as a vector for shRNA to knock down TTK in cells. Briefly, the shRNA targeting TTK was synthesized and ligated into the lentivirus vector PLKO.1. The constructed plasmid and the packaging plasmid were co-transfected into 293T cells, the cell supernatant was collected, and the virus was purified by ultracentrifugation. The lentivirus was added to the cells, and the cells were screened with puromycin 3 days later, and TTK could be stably knocked down after 2 weeks.
Quantificational real-time polymerase chain reaction
Total RNA was extracted using Trizol reagents (15596018, Thermo, Waltham, USA) from cells according to the manufacturer’s instructions. The cDNA was synthesized using the First Strand cDNA Synthesis Kit (K1681, Thermo, Waltham, USA) with 3 µg of total RNA. Relative expression was analyzed using FastStart Universal SYBR Green Master (ROX) (04913949001, Roche, Basel, Switzerland). Primers: GAPDH (control reference gene), forward: CTCCTGCACCACCAACTGCT, reverse, GGGCCATCCACAGTCTTCTG [25]; TTK, forward, TCCCCAGCGCAGCTTTCTGTAGA, reverse, CCAGTCCTCTGGGTTGTTTGCCAT. The relative expression was normalized using GAPDH and calculated using the 2−∆∆Cq method [26].
Xenograft tumor model
All mice experiments were approved by our hospital. Eight-week-old female nude-BABL/c mice were purchased from Beijing Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China), and they were housed in the SPF animal room (4 in each group). 5 × 106 control cells and shTTK cells, resuspended in 100 µl of PBS, were subcutaneously injected into the mouse groin. Tumor growth was observed every 4 days and the size of the tumor was measured to calculate the tumor volume using the formula: Tumor volume = (length × width2)/2 [27]. All mice were sacrificed and xenograft tumors were removed and photographed 31 days after tumor cell injections. The mice were killed by cervical dislocation.
Ethics
All applicable international, national, and/or institutional guidelines for the care and use of human specimens and animals were followed. The animal study was carried out in accordance with the guidelines approved by the Animal Experimentation Ethics Committee of Tianjin Medical University Cancer Institute and Hospital. The protocol was approved by the Committee, and all efforts were made to minimize suffering.
Statistical analysis
GraphPad 5.0 was used for statistical analysis in this study. All data were represented as mean ± standard deviation. Student’s t-test was used for statistical comparisons. Meanwhile, the correlation analysis between clinical pathological features and TTK expression was performed through χ2 analysis. Asteric indicates p < 0.05 and represents significance.
Results
Phosphotyrosine picked threonine kinase showed abnormally high expression in human osteosarcoma tissues
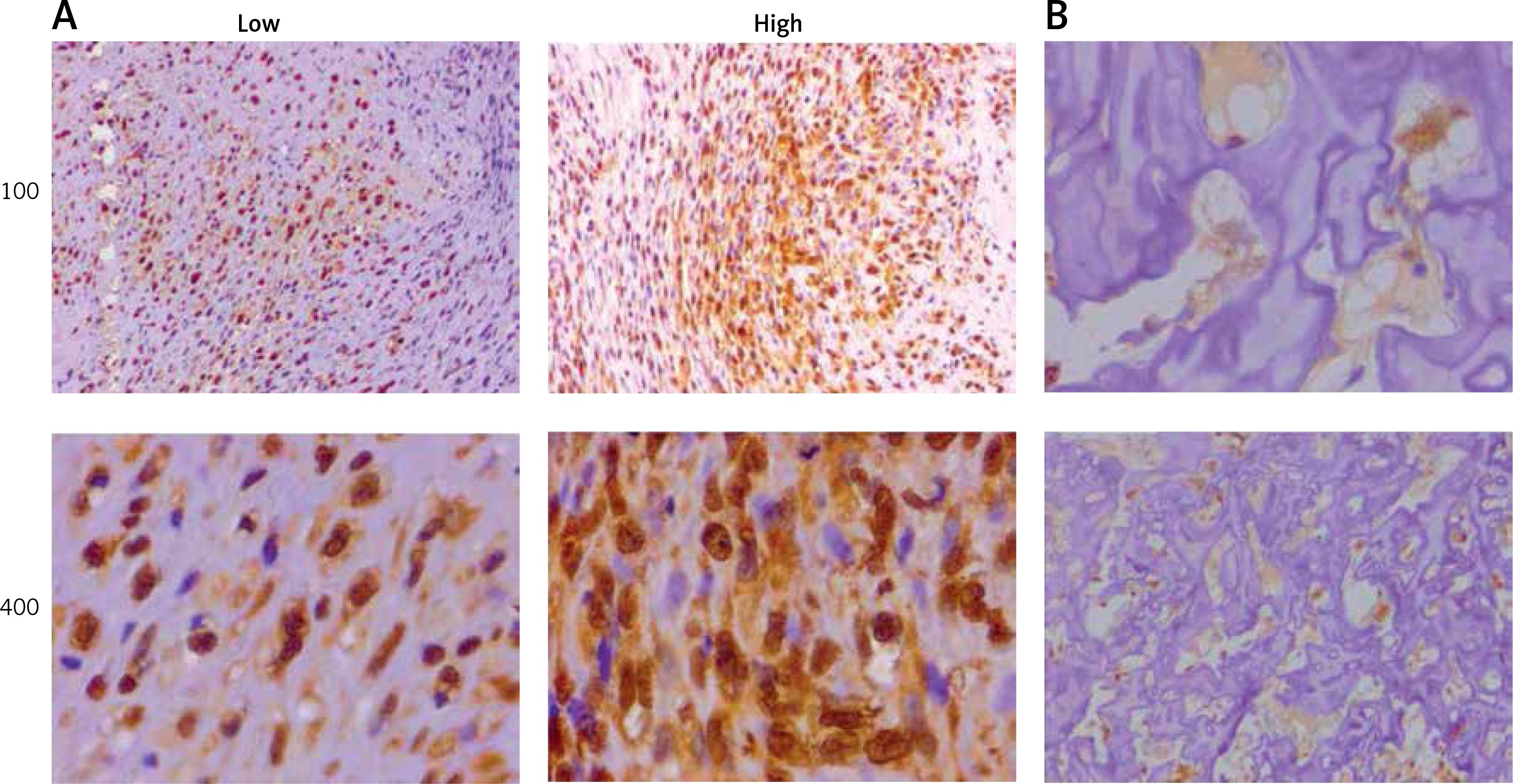
As an oncogene, TTK has been previous reported in various types of tumors. We collected 74 specimens of human OS and tumor information (Table I). By IHC staining, we found that the expression levels of TTK was significantly different in tumor tissues and the corresponding non-tumor tissues (Figure 1 A), and the expression of TTK was significantly correlated with clinical pathological features including tumor size and grade (Table I). As a comparison, TTK showed no or low expression in adjacent normal tissues (Figure 1 B). In conclusion, we observed high expression of TTK in human OS cells and the expression of TTK is correlated with the staging and grading of tumors.
Table I
Relationships of phosphotyrosine picked threonine kinase and clinicopathological characteristics in 74 patients with osteosarcoma
Figure 1
Phosphotyrosine picked threonine kinase (TTK) showed abnormally high expression in human osteosarcoma (OS) tissues. A – To detect TTK expression levels in human OS tissues, immunohistochemical (IHC) assays were performed, and representative photographs are shown (100× and 400× magnification, respectively). B – Results of IHC assays showing the expression levels of TTK in adjacent non-tumor tissues (100× and 400× magnification, respectively)
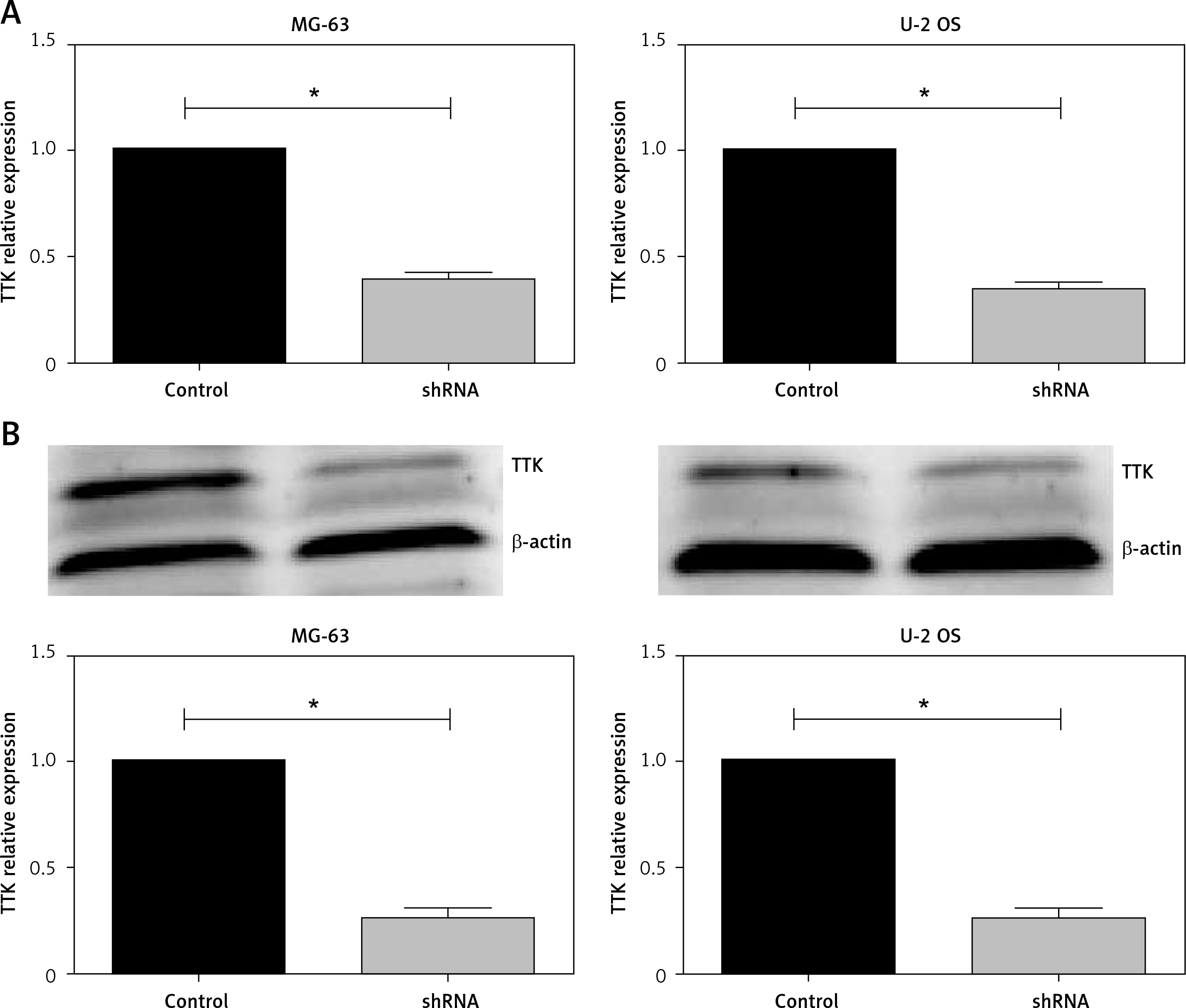
Depletion of phosphotyrosine picked threonine kinase in MG-63 and U-2 OS cells
We then evaluated the possible effect of TTK on proliferation of OS cells in vitro. First, we used lentivirus vector shRNA plasmids to deplete TTK expression in 2 types of OS cell lines, MG-63 and U-2 OS. Detecting the mRNA expression levels of TTK in MG-63 and U-2 OS cells, we found that TTK was significantly decreased when adding shRNA to cells (Figure 2 A), which was further confirmed by Immunoblot assays (Figure 2 B).
Figure 2
Knockdown of phosphotyrosine picked threonine kinase (TTK) using shRNA plasmids targeted TTK in MG-63 and U-2 osteosarcoma (OS) cell lines. A – Expression levels of TTK mRNA in in control cells and knockdown cell in MG-63 (left) and U-2 OS (right). B – Detecting the expression levels of TTK protein in control TTK depletion cells in MG-63 (left) and U-2 OS (right) using western blotting. Quantified using ImageJ software (down)
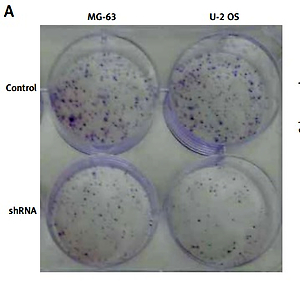
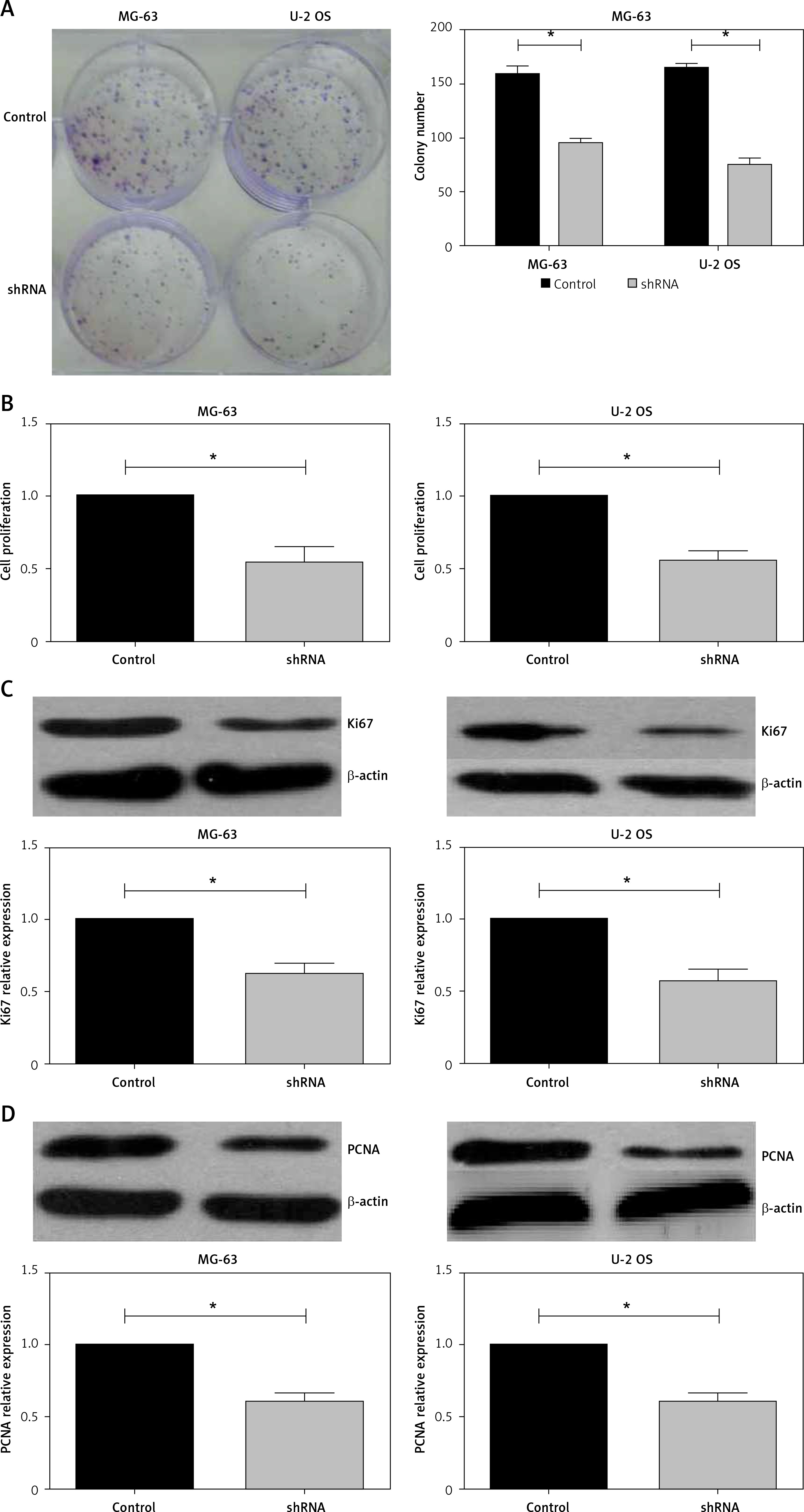
Phosphotyrosine picked threonine kinase ablation blocked cell proliferation in human osteosarcoma cell lines
In human tumor specimens, we previous demonstrated that TTK expression was correlated with tumor size, and therefore we established a cell line that stably depleted TTK by lentivirus vector. Liu et al. reported that TTK promotes cell proliferation and migration in HCC cells [19]. Then, we observed whether TTK ablation affects cell proliferation in OS cells. In the cell colony formation assay, knockdown of TTK in MG-63 and U-2 OS cells decreased colony numbers (Figure 3 A), similarly to the results of cell proliferation assay using CCK-8 reagent (Figure 3 B). Importantly, protein expression of Ki67 and PCNA was detected in control and TTK depleted cells, showing that they had lower expression levels in TTK depletion cells.
Figure 3
Knockdown of phosphotyrosine picked threonine kinase (TTK) weakens the proliferation of osteosarcoma cells. A – Colony formations in MG-63 and U-2 osteosarcoma (OS) cell lines in control and knockdown cells. B – Detecting cell proliferation using cell counting kit-8 assay in MG-63 and U-2 OS cell lines in control and TTK depletion cells. C – Expression level of Ki67 and proliferating cell nuclear antigen in MG-63 and U-2 OS cell lines in control and TTK depletion cells D – Expression level of Ki67 and proliferating cell nuclear antigen in MG-63 and U-2 OS cell lines in control and TTK depletion cells
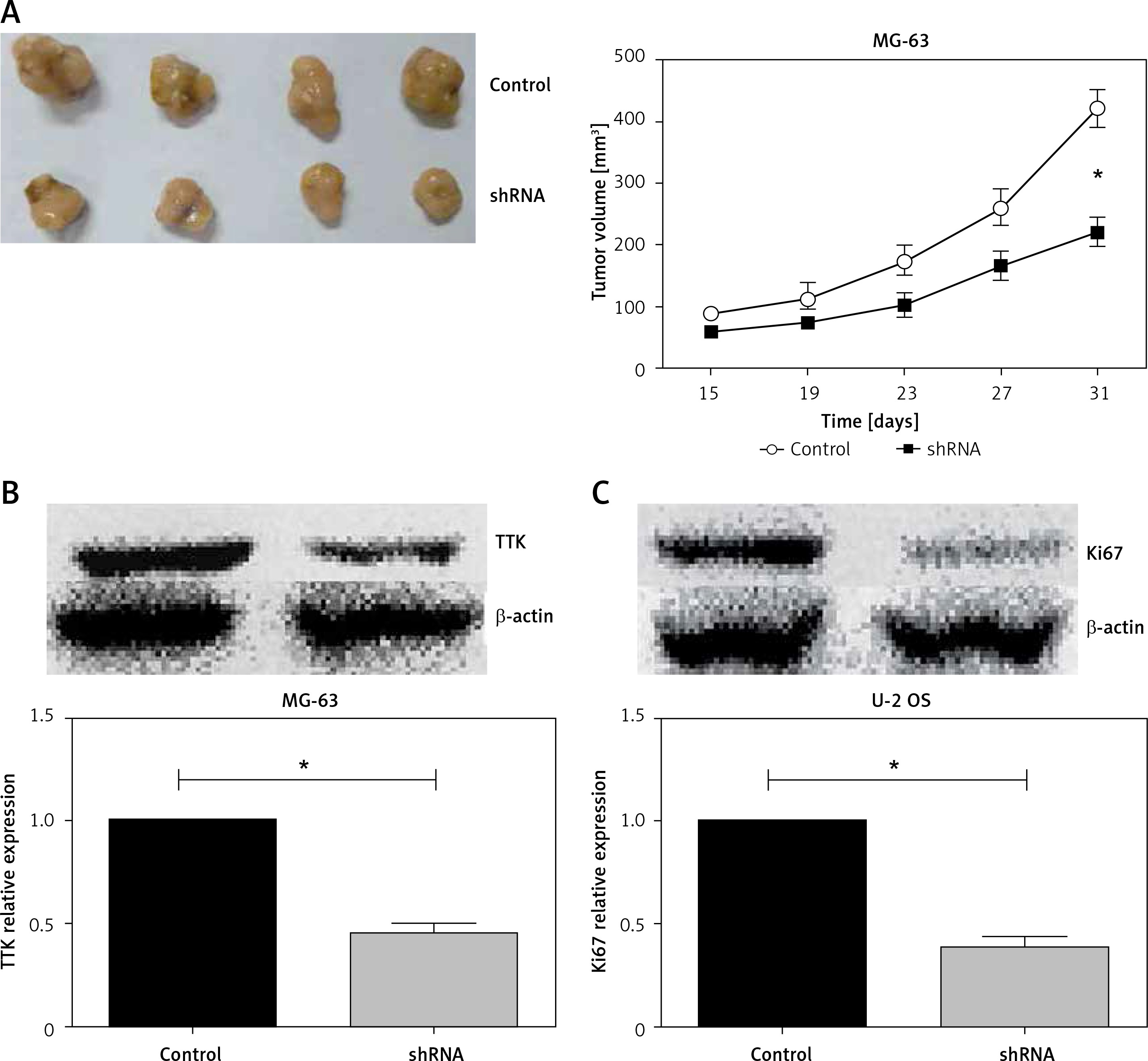
Knockdown of phosphotyrosine picked threonine kinase suppressed tumor growth of osteosarcoma cells in mice
Above, we verified that knockdown of TTK attenuated cell proliferation in vitro. We then evaluated the effects of TTK on tumor growth in vivo using a xenograft tumor model. 5 × 106 cells were inoculated subcutaneously in nude mice, tumor growth was observed every 4 days, tumor size was measured, and the volume of the tumor was calculated. 31 days after the inoculation, the animals were sacrificed and the tumors were removed. As shown in Figure 4 A, tumors were significantly reduced after the depletion of TTK. The expression of TTK in xenograft tumors was detected using Western blotting; as previously described, the expression of TTK was significantly reduced in tumors from TTK depletion groups. Ki67 responded to cell proliferation, and we noted that expression of Ki67 in tumors was also significantly attenuated in the TTK depletion groups. In summary, through the in vivo and in vitro experiments, we found that knocking down TTK attenuated tumor growth.
Discussion
In previous studies, TTK was reported to play a regulatory role in cell mitosis [28–31] and high expression of TTK increases genomic instability and promotes cancer cell proliferation [19]. In a variety of cancers, the expression of TTK is inversely related to the prognosis of the patients, and TTK promotes proliferation and migration in cancer cells in vivo and in vitro. But there is no research indicating the possible role of TTK in human OS progression.
In this research, we collected patients’ information about TTK expression levels and OS stage, confirmed the clinical correlation between high expression of TTK and tumor size and clinical stage. Similar observations have been reported in thyroid papillary carcinoma, breast cancer, gastric cancer, bronchogenic carcinoma, and lung cancers [20]. In breast cancer, King et al. examined a publicly available database and reached a similar conclusion, high expression of TTK with adverse features [22]. In the HCC database, Liu et al. found that TTK was overexpressed in 78% of patients with hepatocellular carcinoma, and high expression positively correlated with tumor size and tumor thrombus [19].
We used shRNA plasmids to deplete TTK expression in human OS cells, MG-63 and U-2 OS, using shRNA in lentivirus and selected stable cells using puromycin. We verified that the cell proliferation was decreased in knockdown TTK cells using colony formation assay and Ki67, a PCNA protein marker in vitro. In the animal experiment, similar results were verified. King et al. showed that TTK leads to low expression of KLF5, resulting in downregulation of miR-200, upregulation of miR-21, and induction of mesenchymal epithelial transition in triple negative breast cancer [22]. The study by Yu et al. showed that TTK promotes cell proliferation and migration via AKT and MDM2 [23] pathway in hepatocellular cells [19].
Several studies have shown that TTK promotes proliferation and migration of tumor cells, so TTK has received high attention as a potential clinical therapeutic target [14], and inhibition of TTK enhanced the efficacy of docetaxel in breast cancer [20]. The inhibition of TTK significantly reduced survival of TNBC cells and showed synergy with docetaxel in cancer cells and a mouse model of human TNBC [20]. Compelling evidence indicated that patients with TNBC with CDKN2A or RB mutation benefit from inhibition [32].
There are some limitations in the study such as the single-center small sample clinical results, the lack of in-depth research on mechanisms, and the lack of conducted studies on over-expression of TTK plasmids. Moreover, we lacked the data on survival of the in vivo model and patients owing to many reasons, but we can provide the data in the next step of the future study. And we can see the published clinical articles [33–37], whose methods can be referenced by us in the future.
In conclusion, this study showed that knockdown of TTK inhibits cell proliferation in human OS cells in vivo and in vitro. However, the precise molecular mechanisms underlying TTK promoting the progression of OS need further study.