Current issue
Archive
Manuscripts accepted
About the Journal
Editorial office
Editorial board
Section Editors
Abstracting and indexing
Subscription
Contact
Ethical standards and procedures
Most read articles
Instructions for authors
Article Processing Charge (APC)
Regulations of paying article processing charge (APC)
CLINICAL RESEARCH
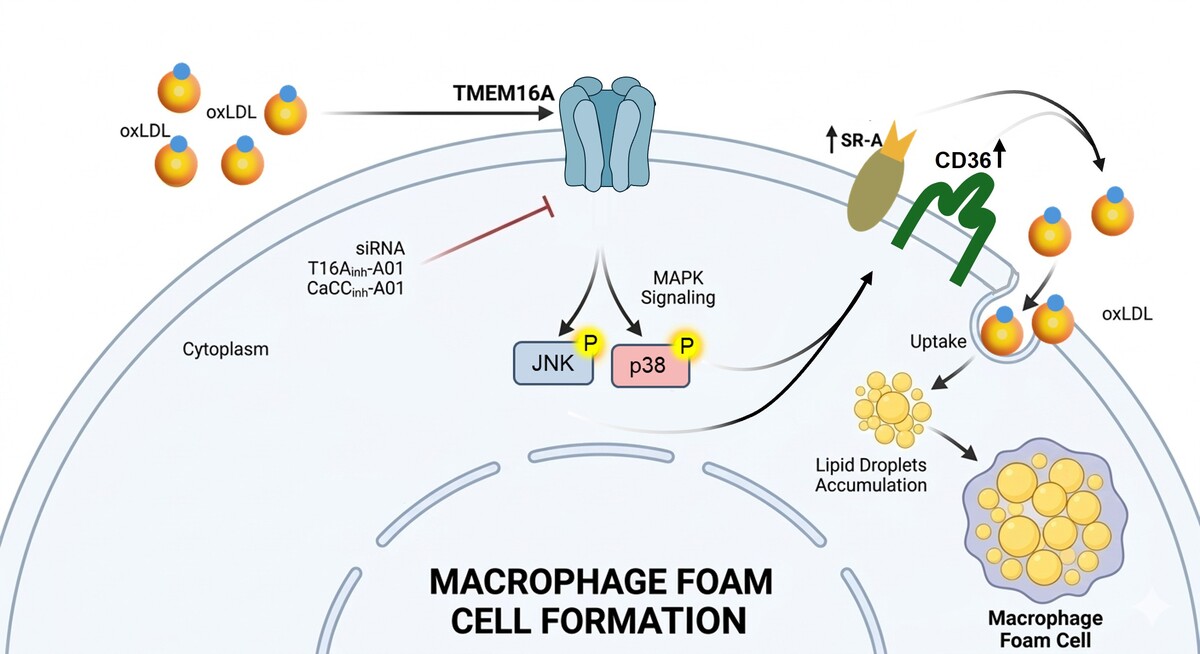
TMEM16A contributes to oxLDL-induced macrophage foam cell formation via the p38/JNK MAPK signaling pathways
1
Department of Pharmacology, Cardiac and Cerebral Vascular Research Center, Zhongshan School of Medicine, Sun Yat-Sen University, Guangzhou, China
2
Department of Cardiovascular Surgery of the First Affiliated Hospital and Institute for Cardiovascular Science, Suzhou Medical College, Soochow University, Suzhou, China
Submission date: 2026-01-30
Final revision date: 2026-03-10
Acceptance date: 2026-04-09
Online publication date: 2026-06-30
Corresponding author
Yanhua Du
Department of Pharmacology Zhongshan School of Medicine Sun Yat-Sen University 74 Zhongshan 2nd Road Guangzhou, Guangdong 510080, China Phone: 86-20-87334787
Department of Pharmacology Zhongshan School of Medicine Sun Yat-Sen University 74 Zhongshan 2nd Road Guangzhou, Guangdong 510080, China Phone: 86-20-87334787
Guozheng Liang
Department of Pharmacology Zhongshan School of Medicine Sun Yat-Sen University 74 Zhongshan 2nd Road Guangzhou, Guangdong 510080, China Phone: 86-20-87334787
Department of Pharmacology Zhongshan School of Medicine Sun Yat-Sen University 74 Zhongshan 2nd Road Guangzhou, Guangdong 510080, China Phone: 86-20-87334787
KEYWORDS
TOPICS
ABSTRACT
Introduction:
Atherosclerotic cardiovascular disease remains the leading cause of global mortality, characterized by lipid accumulation and inflammatory cell infiltration within the arterial wall. A critical early event in atherogenesis is the formation of macrophage foam cells driven by excessive uptake of oxidized low-density lipoprotein (oxLDL). While cations such as Ca²+ have been extensively studied in this context, the role of Cl- as a regulatory anion in foam cell formation remains poorly understood. The calcium-activated chloride channel TMEM16A plays an important role in many physiological processes. However, its role in macrophages remains unclear.
Material and methods:
Macrophage cells were treated with oxLDL to induce foam cell formation. TMEM16A expression was modulated by siRNA-mediated knockdown or cDNA-driven overexpression, and its activity was pharmacologically inhibited using T16Ainh-A01 and CaCCinh-A01. Expression of scavenger receptor A (SR-A) and CD36, phosphorylation level of JNK, and p38 MAPK signaling pathways were analyzed by Western blotting to elucidate underlying mechanisms.
Results:
OxLDL treatment significantly increased TMEM16A expression in RAW264.7 and THP-1 macrophages. Genetic silencing or pharmacological inhibition of TMEM16A markedly attenuated oxLDL-induced lipid accumulation and foam cell formation, whereas TMEM16A overexpression promoted oxLDL accumulation in macrophages. Mechanistically, TMEM16A deficiency significantly reduced SR-A and CD36 expression, which was associated with decreased phosphorylation of JNK and p38 MAPK, indicating that TMEM16A regulates SR-A and CD36 through a MAPK-dependent signaling pathway.
Conclusions:
These findings demonstrate that TMEM16A plays a critical role in macrophage derived foam cell formation by promoting oxLDL uptake through activation of the JNK/p38–SR-A/CD36 axis. Targeting TMEM16A could be a promising therapeutic strategy for atherosclerosis.
Atherosclerotic cardiovascular disease remains the leading cause of global mortality, characterized by lipid accumulation and inflammatory cell infiltration within the arterial wall. A critical early event in atherogenesis is the formation of macrophage foam cells driven by excessive uptake of oxidized low-density lipoprotein (oxLDL). While cations such as Ca²+ have been extensively studied in this context, the role of Cl- as a regulatory anion in foam cell formation remains poorly understood. The calcium-activated chloride channel TMEM16A plays an important role in many physiological processes. However, its role in macrophages remains unclear.
Material and methods:
Macrophage cells were treated with oxLDL to induce foam cell formation. TMEM16A expression was modulated by siRNA-mediated knockdown or cDNA-driven overexpression, and its activity was pharmacologically inhibited using T16Ainh-A01 and CaCCinh-A01. Expression of scavenger receptor A (SR-A) and CD36, phosphorylation level of JNK, and p38 MAPK signaling pathways were analyzed by Western blotting to elucidate underlying mechanisms.
Results:
OxLDL treatment significantly increased TMEM16A expression in RAW264.7 and THP-1 macrophages. Genetic silencing or pharmacological inhibition of TMEM16A markedly attenuated oxLDL-induced lipid accumulation and foam cell formation, whereas TMEM16A overexpression promoted oxLDL accumulation in macrophages. Mechanistically, TMEM16A deficiency significantly reduced SR-A and CD36 expression, which was associated with decreased phosphorylation of JNK and p38 MAPK, indicating that TMEM16A regulates SR-A and CD36 through a MAPK-dependent signaling pathway.
Conclusions:
These findings demonstrate that TMEM16A plays a critical role in macrophage derived foam cell formation by promoting oxLDL uptake through activation of the JNK/p38–SR-A/CD36 axis. Targeting TMEM16A could be a promising therapeutic strategy for atherosclerosis.
REFERENCES (48)
1.
Sarraju A, Nissen SE. Atherosclerotic plaque stabilization and regression: a review of clinical evidence. Nat Rev Cardiol 2024; 21: 487-97.
2.
Banach M, Surma S, Toth PP. 2023: The year in cardiovascular disease – the year of new and prospective lipid lowering therapies. Can we render dyslipidemia a rare disease by 2024? Arch Med Sci 2023; 19: 1602-15.
3.
Takaoka M, Zhao X, Lim HY, et al. Early intermittent hyperlipidaemia alters tissue macrophages to fuel atherosclerosis. Nature 2024; 634: 457-65.
4.
Ai J, Tang X, Zhou Y, et al. New insights into foam cells in atherosclerosis. Cardiovasc Res 2025; 121: 2334-46.
5.
Mukherjee P, Rahaman SG, Goswami R, et al. Role of mechanosensitive channels/receptors in atherosclerosis. Am J Physiol Cell Physiol 2022; 322: C927-38.
6.
Lan Y, Lu J, Zhang S, et al. Piezo1-mediated mechanotransduction contributes to disturbed flow-induced atherosclerotic endothelial inflammation. J Am Heart Assoc 2024; 13: e035558.
7.
Atcha H, Kulkarni D, Meli VS, et al. Piezo1-mediated mechanotransduction enhances macrophage oxidized low-density lipoprotein uptake and atherogenesis. PNAS Nexus 2024; 3: pgae436.
8.
Zhang Y, Ma K, Fang X, et al. Targeting ion homeostasis in metabolic diseases: Molecular mechanisms and targeted therapies. Pharmacol Res 2025; 212: 107579.
9.
Azlan NFM, Zhang J. Role of the cation-chloride-cotransporters in cardiovascular disease. Cells 2020; 9: 2293.
10.
Wang M, Yang H, Zheng LY, et al. Downregulation of TMEM16A calcium-activated chloride channel contributes to cerebrovascular remodeling during hypertension by promoting basilar smooth muscle cell proliferation. Circulation 2012; 125: 697-707.
11.
Raut SK, Singh K, Sanghvi S, et al. Chloride ions in health and disease. Biosci Rep 2024; 44: BSR20240029.
12.
Wu QQ, Liu XY, Xiong LX, et al. Reduction of intracellular chloride concentration promotes foam cell formation. Circ J 2016; 80: 1024-33.
13.
Li X, Du YX, Yu CL, Niu N. Ion channels in macrophages: implications for disease progression. Int Immunopharmacol 2025; 144: 113628.
14.
Zeng XL, Sun L, Zheng HQ, et al. Smooth muscle-specific TMEM16A expression protects against angiotensin II-induced cerebrovascular remodeling via suppressing extracellular matrix deposition. J Mol Cell Cardiol 2019; 134: 131-43.
15.
Al-Hosni R, Kaye R, Choi CS, Tammaro P. The TMEM16A channel as a potential therapeutic target in vascular disease. Curr Opin Nephrol Hypertens 2024; 33: 161-9.
16.
Zhang X, Zheng B, Yang Z, et al. TMEM16A and myocardin form a positive feedback loop that is disrupted by KLF5 during Ang II–induced vascular remodeling. Hypertension 2015; 66: 412-21.
17.
Lv XF, Zhang YJ, Liu X, et al. TMEM16A ameliorates vascular remodeling by suppressing autophagy via inhibiting Bcl-2-p62 complex formation. Theranostics 2020; 10: 3980-93.
18.
Tao J, Liu CZ, Yang J, et al. ClC-3 deficiency prevents atherosclerotic lesion development in ApoE-/- mice. J Mol Cell Cardiol 2015; 87: 237-47.
19.
Hong L, Xie ZZ, Du YH, et al. Alteration of volume-regulated chloride channel during macrophage-derived foam cell formation in atherosclerosis. Atherosclerosis 2011; 216: 59-66.
20.
Genovese M, Buccirossi M, Guidone D, et al. Analysis of inhibitors of the anoctamin-1 chloride channel (transmembrane member 16A, TMEM16A) reveals indirect mechanisms involving alterations in calcium signalling. Br J Pharmacol 2023; 180: 775-85.
21.
Bradley E, Fedigan S, Webb T, et al. Pharmacological characterization of TMEM16A currents. Channels 2014; 8: 308-20.
22.
Dejager S, Mietus-Synder M, Pitas RE. Oxidized low density lipoproteins bind to the scavenger receptor expressed by rabbit smooth muscle cells and macrophages. Arterioscler Thromb 1993; 13: 371-8.
23.
Lian TW, Wang L, Lo YH, Huang IJ, Wu MJ. Fisetin, morin and myricetin attenuate CD36 expression and oxLDL uptake in U937-derived macrophages. Biochim Biophys Acta 2008; 1781: 601-9.
24.
Kunjathoor VV, Febbraio M, Podrez EA, et al. Scavenger receptors class A-I/II and CD36 are the principal receptors responsible for the uptake of modified low density lipoprotein leading to lipid loading in macrophages.
26.
Liu Q, Fan J, Bai J, et al. IL-34 promotes foam cell formation by enhancing CD36 expression through p38 MAPK pathway. Sci Rep 2018; 8: 17347.
27.
Yin R, Dong Y, Li H. PPAR phosphorylation mediated by JNK MAPK: a potential role in macrophage-derived foam cell formation. Acta Pharmacol Sin 2006; 27: 1146-52.
28.
Mazzone A, Eisenman ST, Strege PR, et al. Inhibition of cell proliferation by a selective inhibitor of the Ca(2+)-activated Cl(-) channel, Ano1. Biochem Biophys Res Commun 2012; 427: 248-53.
29.
Bill A, Hall ML, Borawski J, et al. Small molecule-facilitated degradation of ANO1 protein: a new targeting approach for anticancer therapeutics. J Biol Chem 2014; 289: 11029-41.
30.
Weir HK, Anderson RN, Coleman King SM, et al. Heart disease and cancer deaths – trends and projections in the United States, 1969–2020. Prev Chronic Dis 2016; 13: E157.
31.
Ceasovschih A, Banjanin N, Bednarek A, et al. Atherosclerotic features in patients with heart failure. Arch Med Sci 2025; 21: 1107-29.
32.
Toth PP, Banach M. 2025: The year in cardiovascular disease – the year of triglyceride lowering therapies. Can we effectively reduce triglyceride-related residual cardiovascular disease and pancreatitis risk? Arch Med Sci 2025; 21: 2229-45.
33.
Tabas I, Bornfeldt KE. Macrophage phenotype and function in different stages of atherosclerosis. Circ Res 2016; 118: 653-67.
34.
Duan DD. The ClC-3 chloride channels in cardiovascular disease. Acta Pharmacol Sin 2011; 32: 675-84.
35.
Goto K, Kitazono T. Chloride ions, vascular function and hypertension. Biomedicines 2022; 10: 2316.
36.
Oh J, Riek AE, Weng S, et al. Endoplasmic reticulum stress controls M2 macrophage differentiation and foam cell formation. J Biol Chem 2012; 287: 11629-41.
37.
Ricote M, Li AC, Willson TM, Kelly CJ, Glass C K. The peroxisome proliferator-activated receptor- is a negative regulator of macrophage activation. Nature 1998; 391: 79-82.
38.
Allawzi AM, Vang A, Clements RT, et al. Activation of anoctamin-1 limits pulmonary endothelial cell proliferation via p38–mitogen-activated protein kinase–dependent apoptosis. Am J Respir Cell Mol Biol 2018; 58: 658-67.
39.
Liu D, Wang K, Su D, et al. TMEM16A regulates pulmonary arterial smooth muscle cells proliferation via p38MAPK/ERK pathway in high pulmonary blood flow-induced pulmonary arterial hypertension. J Vasc Res 2020; 58: 27-37.
40.
Sun W, Guo S, Li Y, et al. Anoctamin 1 controls bone resorption by coupling Cl- channel activation with RANKL-RANK signaling transduction. Nat Commun 2022; 13: 2899.
41.
Zhang Y, Zheng HQ, Chen BY, et al. WNK1 is required for proliferation induced by hypotonic challenge in rat vascular smooth muscle cells. Acta Pharmacol Sin 2018; 39: 35-47.
42.
Zheng H, Li X, Zeng X, et al. TMEM16A inhibits angiotensin II-induced basilar artery smooth muscle cell migration in a WNK1-dependent manner. Acta Pharm Sin B 2021; 11: 3994-4007.
43.
Garrud TAC, Bell B, Mata-Daboin A, et al. WNK kinase is a vasoactive chloride sensor in endothelial cells. Proc Natl Acad Sci USA 2024; 121: e2322135121.
44.
Mayes-Hopfinger L, Enache A, Xie J, et al. Chloride sensing by WNK1 regulates NLRP3 inflammasome activation and pyroptosis. Nat Commun 2021; 12: 4546.
45.
Ma MM, Gao M, Guo KM, et al. TMEM16A contributes to endothelial dysfunction by facilitating Nox2 NADPH oxidase–derived reactive oxygen species generation in hypertension. Hypertension 2017; 69: 892-901.
46.
Kimura Y, Yanagida T, Onda A, et al. Soluble uric acid promotes atherosclerosis via AMPK (AMP-activated protein kinase)-mediated inflammation. Arterioscler Thromb Vasc Biol 2020; 40: 570-82.
47.
Niu SW, Hung CC, Lin HYH, et al. Reduced incidence of stroke in patients with gout using benzbromarone. J Pers Med 2022; 12: 28.
48.
Al-Hosni R, Ilkan Z, Agostinelli E, Tammaro P. The pharmacology of the TMEM16A channel: therapeutic opportunities. Trends Pharm Sci 2022; 43: 712-25.
Share
RELATED ARTICLE
| eISSN: | 1896-9151 |
| ISSN: | 1734-1922 |
We process personal data collected when visiting the website. The function of obtaining information about users and their behavior is carried out by voluntarily entered information in forms and saving cookies in end devices. Data, including cookies, are used to provide services, improve the user experience and to analyze the traffic in accordance with the Privacy policy. Data are also collected and processed by Google Analytics tool (more).
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.