Introduction
Metabolic dysfunction-associated steatotic liver disease (MASLD) is a condition characterized by the accumulation of fat in the liver as a consequence of metabolic disturbances, such as obesity, insulin resistance (IR), and dyslipidemia. MASLD serves as an umbrella term that encompasses a broad spectrum of pathological liver changes – from simple hepatic steatosis to steatohepatitis associated with metabolic dysfunction, which can lead to fibrosis and eventually progress to cirrhosis or hepatocellular carcinoma (HCC), while simultaneously increasing the risk of cardiovascular diseases (CVDs) and extrahepatic malignancies [1].
Given the complex pathophysiology and multisystemic implications of MASLD, there is a growing need for integrative strategies that facilitate early diagnosis, disease monitoring, and targeted interventions. The novelty of this review lies in its comprehensive approach that brings together biomarkers of oxidative stress, inflammation, and genetic/epigenetic regulation within a clinically oriented multi-omics and an artificial intelligence (AI)-guided framework. Moreover, we highlight recent therapeutic advances – including the Food and Drug Administration (FDA) approval of resmetirom in 2024 and other promising agents currently in phase 2/3 trials – providing a timely and translationally relevant overview that bridges current scientific evidence with emerging clinical applications in MASLD.
Epidemiology, risk factors, and natural course of the disease
The prevalence of MASLD (previously known as non-alcoholic fatty liver disease – NAFLD) has been extensively studied worldwide over the past decades. MASLD is recognized as the most common liver disease globally, affecting approximately 30% of the population, with its prevalence increasing from 22% to 37% over the past two decades [2]. MASLD is particularly prevalent among individuals with metabolic dysfunction, with a reported prevalence of up to 68% among patients with type 2 diabetes mellitus (T2DM), while obese individuals have more than a threefold elevated risk of developing this condition [3, 4]. The global prevalence of MASLD increases in parallel with obesity rates, and a recent meta-analysis determined a prevalence of 75.27% in the obese population [5]. CVDs are the leading cause of death in patients with MASLD, whereas liver-related mortality is confined to patients with advanced fibrosis, with the risk rising exponentially as the fibrosis stage increases [6]. Metabolic dysfunction-associated steatohepatitis (MASH) is a more dangerous form of MASLD, characterized by histological findings of inflammation in the liver parenchyma, hepatocyte ballooning with increased hepatocyte necrosis, and an elevated risk of fibrosis progression [7]. A meta-analysis conducted by Lim et al., which included 22 global observational studies encompassing a total of 379,801 patients, demonstrated that the overall prevalence of MASLD was 39.22% (95% CI: 30.96% to 48.15%) [8]. Approximately 10% of patients with MASLD, that is, 2–5% of the general population in the USA, have MASH [9]. In patients without an indication for liver biopsy, the prevalence of MASH is 7% [10]. Liver cirrhosis, a frequent complication resulting from MASH, is not only the leading cause of HCC but also the second most common indication for liver transplantation in the USA [11]. Existing data on MASLD prevalence indicate higher rates in Western countries compared to Eastern nations, although a marked upward trend has been observed in the latter over the past decade [12]. In the coming years, an increase in the number of MASLD cases, along with its associated complications, including HCC, is anticipated [13]. The MASLD epidemic, with its high prevalence in the general population, is attributed to numerous factors, the most important being the availability of high-calorie foods and reduced levels of physical activity. Globalization and the adoption of a Western-style diet in developing countries, combined with a sedentary lifestyle and rising obesity rates, contribute significantly to the high prevalence of MASLD [14].
The term “NAFLD” was first introduced in 1980 to characterize the clinical presentation of steatohepatitis, a severe liver injury characterized by lobular inflammation, steatosis, and ballooning degeneration of hepatocytes in patients with obesity and other metabolic disorders who did not consume alcohol [15]. However, more than four decades later, in light of significant epidemiological changes driven by shifts in lifestyle and new dietary habits worldwide, questions began to emerge regarding the adequacy of this terminology. In response to these changes, in 2020 an international group of experts proposed a new classification, termed metabolic dysfunction-associated fatty liver disease (MAFLD) [16, 17]. The introduction of this term was met with criticism from parts of the professional community, who expressed concern that the new nomenclature might obscure clinically important differences in the pathophysiology and natural history of liver injury between metabolic and alcoholic etiologies. This concern arose because the diagnosis of MAFLD required the presence of hepatic steatosis along with obesity, T2DM, or at least two additional metabolic risk factors, without excluding patients with significant alcohol consumption [18]. Given the resistance among experts to this proposal and in order to address these ambiguities, a multidisciplinary panel later recommended replacing the term NAFLD with MASLD [18].
The diagnosis of MASLD is based on the confirmed presence of hepatic steatosis, determined by imaging techniques or biopsy, in combination with cardiometabolic risk factors such as obesity (i.e., elevated body mass index – BMI), hypertension, IR, or dyslipidemia, while excluding significant alcohol consumption [18]. To diagnose hepatic steatosis, it is necessary to demonstrate fat accumulation in more than 5% of hepatocytes on histopathological examination, in the absence of significant alcohol consumption or other causes of liver injury or steatosis, or to detect more than 5.6% fat using proton magnetic resonance spectroscopy (¹H-MRS) [19]. Alcohol and medications are common causes of fat deposition in hepatocytes; therefore, in order to diagnose MASLD, it is essential to exclude other potential causes such as excessive alcohol consumption (defined as > 20 g/day, i.e., > 14 drinks per week for women, or > 30 g/day, i.e., > 21 drinks per week for men) and, via a comprehensive medical history, rule out other chronic liver diseases and the use of various drugs that may lead to steatosis [10, 19]. The consensus of the last multidisciplinary panel also defines a subgroup of patients who meet the MASLD criteria but consume higher amounts of alcohol (210–420 g per week for men and 140–350 g per week for women), categorizing them under metabolic dysfunction and alcohol-associated steatotic liver disease (MetALD) or a combined diagnosis [20].
MASLD generally progresses slowly in both adults and children. However, fibrosis can develop much more rapidly in approximately 20% of patients [7]. Environmental factors, including diet, physical activity levels, endocrine disorders, dysbiosis of the gastrointestinal microbiota, and genetic predisposition, can significantly influence disease development and outcomes [21]. It is estimated that the average time for fibrosis progression per stage in patients with MASLD is approximately 14 years, while in those with MASH, this period is roughly halved to about 7 years. Furthermore, the presence of arterial hypertension may further accelerate fibrosis progression, shortening this interval by an additional 50% [7]. Although HCC is the most severe complication of liver cirrhosis within the MASLD spectrum, it can also develop in patients with MASH even in the absence of overt cirrhosis [22]. Unfortunately, despite its high prevalence, a large number of MASLD cases remain undiagnosed, often leading to a diagnosis of cryptogenic cirrhosis, where MASH is the underlying condition once other etiological factors for chronic liver disease, such as alcohol abuse, viral hepatitis, autoimmune disorders, and other metabolic diseases, have been excluded [23]. These findings underscore the critical need for an accurate and widely accessible non-invasive marker capable of detecting MASLD at its earliest stage, allowing for timely intervention and improved patient outcomes.
Meta-analyses have substantially refined estimates of MASLD prevalence and underscored the role of key metabolic risk factors, particularly in populations with T2DM [4, 24]. These findings provide a foundational epidemiologic context for disease burden and stratification across at-risk groups.
Advancements in omics technologies – especially transcriptomics, lipidomics, and multiomics – have yielded promising insights into the molecular landscape of MASLD. These studies have identified candidate biomarkers and distinct pathophysiologic signatures, which may inform disease classification, risk stratification, and therapeutic targeting [25–29].
In parallel, data from prospective cohort studies have enhanced our understanding of MASLD progression and associated outcomes over time. Such longitudinal evidence is crucial for evaluating biomarker performance in real-world settings and for elucidating the cardiometabolic consequences of hepatic steatosis [30, 31].
Pathogenesis of MASLD
The precise mechanisms underlying MASLD have not yet been fully elucidated, and thus its pathogenesis remains incompletely understood. Disturbances in lipid metabolism represent the primary cause of fatty liver, which in MASLD manifests as the accumulation of triglycerides (TG) in hepatocytes. According to the previously accepted “two-hit theory”, the development of this disease occurs in two stages. The first step in the pathogenesis of MASLD involves the presence of abdominal obesity and/or IR, which collectively contribute to diminished inhibition of lipolysis in visceral adipose tissue. This metabolic dysregulation leads to an increased mobilization and subsequent influx of free fatty acids (FFAs) into the liver, thereby promoting the accumulation of TG and the development of hepatic steatosis. However, while this initial hepatic fat accumulation represents a key early event, it is insufficient on its own to drive disease progression. An additional insult or pathogenic factor, often referred to in the literature as the second hit, is required to trigger the progression from simple steatosis to steatohepatitis (MASH), characterized by hepatocellular injury, inflammation, and varying degrees of fibrosis [32]. Developed steatosis further stimulates de novo lipogenesis, a characteristic feature of MASLD patients, while concurrently reducing lipid efflux in the form of high-density lipoprotein (HDL) particles due to insufficient apoB protein synthesis [33]. The second stage of disease progression involves the transition from simple steatosis to MASH, marked by the onset of inflammation and fibrosis, which occurs in approximately one-third of MASLD patients [32, 34].
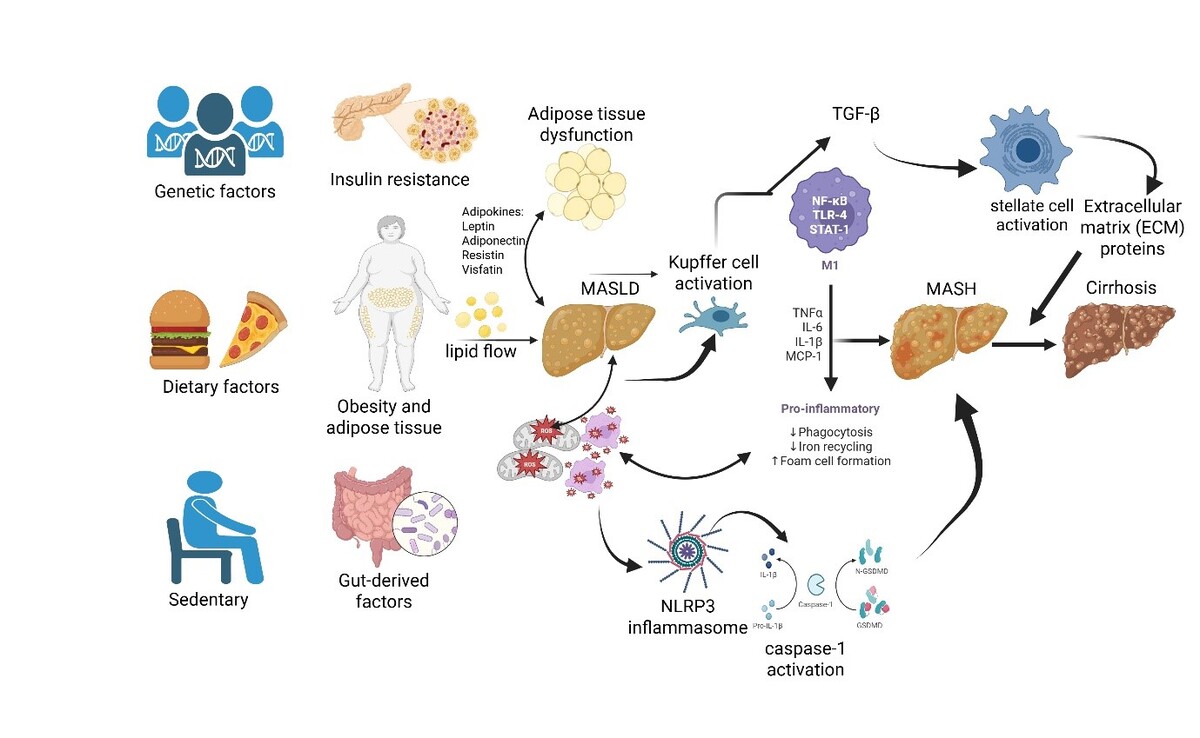
However, over time, this theory has been increasingly revised as new findings emerged and newer models were proposed. It is now known that a wide range of factors influences the development of MASLD, including genetic predisposition [35]. Changes in dietary patterns, alcohol consumption, smoking, and IR also significantly contribute to its pathogenesis and progression [36]. A very high prevalence of MASLD has been observed among obese individuals, which further strengthens the pathophysiological link between obesity and MASLD [37]. In addition to the classical “two-hit theory”, recent findings support a more comprehensive framework known as the “multiple-hit theory”. This model suggests that, beyond TG accumulation and inflammation, additional interrelated mechanisms – such as lipotoxicity, endoplasmic reticulum (ER) stress, mitochondrial dysfunction, and inflammasome activation – collectively contribute to the development and progression of MASLD. These interacting pathways induce hepatocyte injury, apoptosis, and fibrogenesis through oxidative stress and immune activation [38, 39]. The complex interplay of these hits is illustrated in Figure 1, which summarizes the sequential and overlapping mechanisms underlying MASLD pathogenesis.
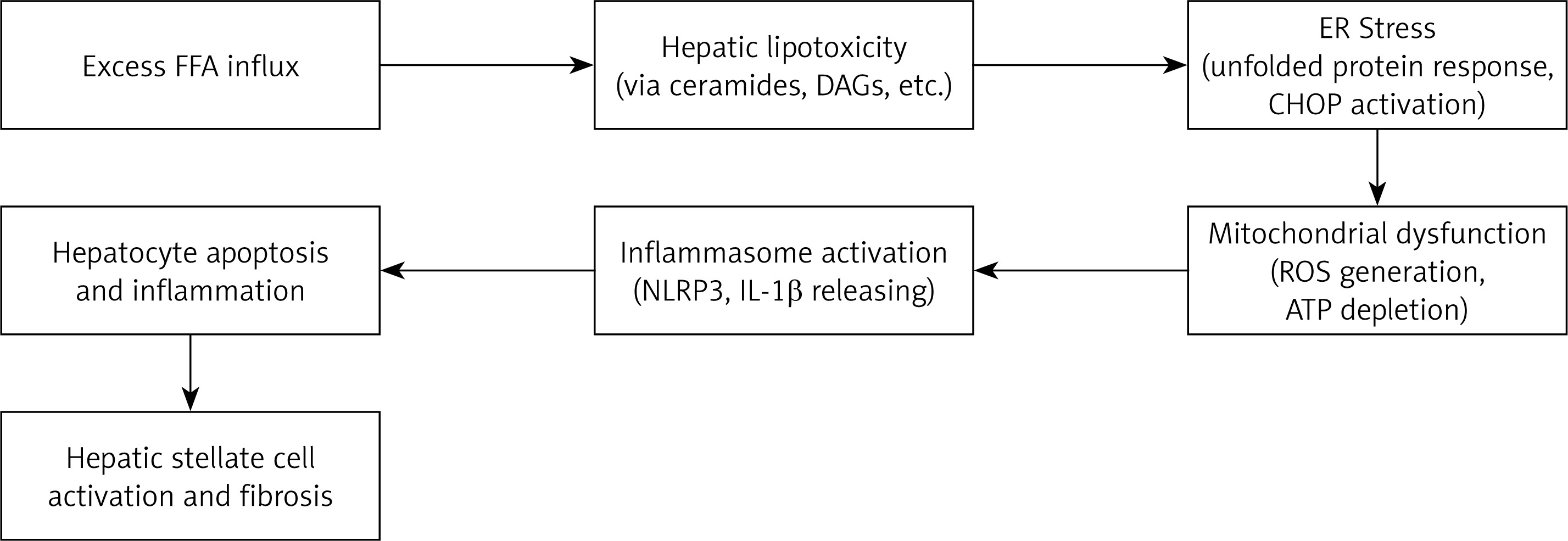
This diagram illustrates the sequential and interacting pathways contributing to MASLD progression. Excess FFA influx initiates hepatic lipotoxicity, which induces endoplasmic reticulum (ER) stress, mitochondrial dysfunction, and inflammasome activation. These insults collectively drive hepatocyte apoptosis, inflammation, and ultimately hepatic stellate cell activation and fibrosis.
Based on these new insights, a theory has been proposed that explains the pathogenesis of MASLD through processes that exceed the normal metabolic capacity of the liver, leading to the excessive accumulation of toxic lipids, hepatocellular stress, injury, and cell death [40]. Recent studies have identified new outcomes of hepatocellular injury, such as ferroptosis, a specific form of iron-dependent cell death that is closely associated with the pathogenesis of MASLD, given the liver’s propensity for iron metabolism and lipid peroxidation. This type of cell death differs from conventional forms, such as autophagy, apoptosis, and necrosis, as it is primarily triggered by the accumulation of lipid peroxides. Biochemically, this process is accompanied by increased lipid peroxidation, elevated levels of reactive oxygen species (ROS), and specific genetic alterations [41]. Additionally, genetic factors may contribute to MASLD outcomes, particularly regarding CVDs, as studies have identified an association between certain patatin-like phospholipase domain-containing protein 3 (PNPLA3) genotypes and cardiovascular disorders [42]. Increased activation of the nucleotide-binding oligomerization domain-like receptors containing pyrin domain 3 (NLRP3) inflammasome has been linked to the development of various conditions, such as atherosclerosis, MASLD, T2DM, and numerous others [39].
Hepatic steatosis can induce oxidative stress, organelle dysfunction, apoptosis, and other pathophysiological disturbances, thereby contributing to the onset and progression of MASLD [43]. Inflammation in adipose tissue is significantly associated with the histological severity of MASLD, with inflammatory processes in visceral adipose tissue potentially preceding and contributing to the progression of the disease to MASH and liver fibrosis [44]. The primary pathophysiological factors underlying MASLD are metabolic dysfunction and atherogenic dyslipidemia [45]. Chronic, low-grade inflammation plays a crucial role in mediating both hepatic and most extrahepatic complications of MASLD. These deranged metabolic processes in MASLD patients, initially mediated by hepatic IR, are further exacerbated by peripheral IR, ultimately resulting in an increased influx of fatty acids into hepatocytes. Visceral adipose tissue serves as a significant source of ROS, proinflammatory adipokines, and cytokines [46]. ROS influence signaling pathways, increase the influx of FFAs into the liver, stimulate de novo lipogenesis, and promote TG deposition, leading to morphological and functional changes in hepatocytes and favoring the onset of apoptosis [47]. Increased fatty acid oxidation does not eliminate hepatic lipid deposits but rather promotes the formation of dysfunctional mitochondria that excessively produce ROS, thereby contributing to oxidative stress and the progression of liver disease [48]. Insulin facilitates the esterification of FFAs and deposition of TG in adipose tissue. With the development of IR, an unfavorable, reverse flux of FFAs toward non-adipose tissues, such as the liver, occurs. Consequently, there is an increased influx of FFAs into the liver, driven both by reduced inhibition of lipolysis and by an increase in de novo synthesis of FFAs [49]. Membrane-bound fatty acid transport proteins (FATPs), especially FATP2 and FATP5, identified in the liver, facilitate the uptake of fatty acids and their availability for hepatic lipid metabolism [50]. In MASLD patients, reduced levels of hepatoprotective adipokines, such as adiponectin, further contribute to hepatic fat accumulation. The increased presence of FFAs in the liver activates tumor necrosis factor-α (TNF-α) production in hepatocytes, which in turn inhibits hepatic adiponectin activity and promotes both MASLD and hepatic IR [51]. Ectopic fat deposition occurs in organs other than the liver, such as the heart, pancreas, and skeletal muscle, leading to cardiometabolic complications in patients with MASLD [18].
The gut microbiome plays a crucial role in maintaining metabolic homeostasis, and its disruption can lead to the development of various metabolic disorders, including MASLD [52]. Studies have confirmed alterations in the composition of the gut microbiota in MASLD patients, particularly among those with advanced stages of the disease, characterized by dysbiosis and the emergence of specific pathobionts [53]. Additionally, changes in microbiota structure are associated with the severity of MASLD, as its diversity decreases in patients with severe disease [54]. Other investigations have demonstrated that gut microbiota analysis has the potential to predict the subsequent development of MASLD [55]. Gut dysbiosis may increase the concentrations of lipopolysaccharides (LPS) in the liver and systemically in MASLD patients [56]. Ethanol, a key metabolite produced by gut microorganisms, is also strongly associated with MASLD [57]. Although experimental models have demonstrated that certain probiotic strains exert beneficial protective and anti-inflammatory effects, promising therapeutic options remain elusive, while dysbiosis, along with the emergence of various pathobionts and gut bacterial metabolites, may contribute to mild hepatic and systemic inflammation [58]. In addition to the fact that excessive caloric intake is recognized as the primary cause of the MASLD epidemic, the choice of food is also critical. Certain dietary components can promote low-grade inflammation in the body through various mechanisms. One such mechanism is the modulation of the gut microbiome, wherein an anti-inflammatory diet may lead to favorable changes in gut microbiota composition [59].
A simplified schematic illustration of the progression from simple hepatic steatosis to cirrhosis is presented in Figure 2.
Figure 2
Schematic illustration of the pathophysiological progression from simple steatosis to cirrhosis in MASLD
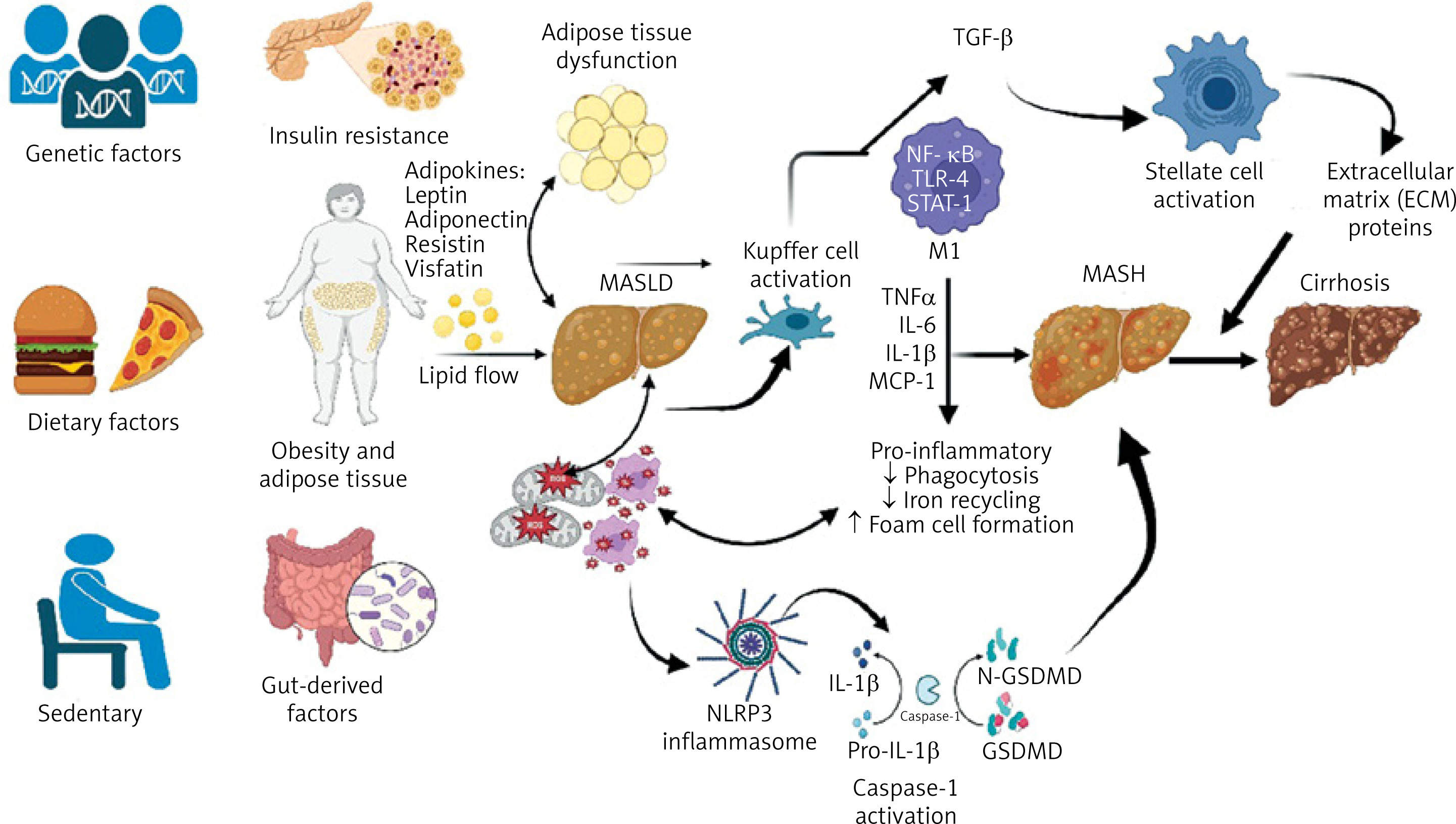
This figure illustrates the multifactorial pathogenesis and sequential progression of metabolic dysfunction-associated steatotic liver disease (MASLD), starting from simple hepatic steatosis and advancing to metabolic dysfunction-associated steatohepatitis (MASH), fibrosis, and ultimately cirrhosis. Genetic predisposition, sedentary lifestyle, dietary factors, and gut-derived elements contribute to obesity, insulin resistance, and adipose tissue dysfunction. These metabolic disturbances lead to hepatic lipid accumulation, Kupffer cell activation, and NLRP3 inflammasome signaling. Subsequent inflammatory cytokine release (TNF-α, IL-6, IL-1β, MCP-1) and activation of hepatic stellate cells promote extracellular matrix deposition and fibrogenesis, driving the progression toward cirrhosis.
MASLD and the risk of cardiovascular disease and malignancies
It is increasingly evident that MASLD is a multisystem disease in which IR and accompanying metabolic dysfunction play a pathogenetic role in both the disease itself and its major complications, including liver-related issues as well as extrahepatic conditions such as CVD, T2DM, chronic kidney disease (CKD), and others [60].
Given the systemic nature of MASLD, this review incorporates interdisciplinary perspectives. IR and T2DM, as core components of endocrine dysfunction, play a central role in the development and progression of MASLD, representing a broad area for ongoing research into novel therapeutic approaches [61]. In parallel, CVD remains the leading cause of death in MASLD patients, underscoring the importance of integrated cardiometabolic risk assessment [62]. Emerging collaborations between hepatologists, endocrinologists, and cardiologists are essential for developing effective, multidisciplinary management strategies [63].
Unhealthy, high-calorie diets rich in saturated fats, refined carbohydrates, sweetened beverages, and fructose, combined with physical inactivity and a sedentary lifestyle, substantially contribute to the onset of obesity and MASLD [64]. MASLD is bidirectionally associated with metabolic disorders related to IR, such as T2DM, polycystic ovary syndrome (PCOS), obesity, and dyslipidemia [65, 66]. IR is also the key pathophysiologic trigger of MASLD, even in the absence of obesity. Hence, lean individuals with metabolic diseases should also be screened for MASLD [63].
MASLD is associated with an approximately 35% increased risk of cardiovascular mortality [67, 68]. Furthermore, a recent meta-analysis of 11 longitudinal cohort studies demonstrated that MASLD is associated with a 1.5-fold higher long-term risk of new-onset heart failure, regardless of the presence of hypertension, T2DM, and other common cardiometabolic risk factors [69]. In general, MASLD increases hepatic IR and stimulates the production of proinflammatory cytokines, leading to the development of various CVDs, including atherosclerotic disease and heart failure. Statin treatment is recommended in all individuals with MAFLD due to their heightened CVD risk. Statins are considered to be safe in patients with MASLD with mildly and moderately increased serum transaminases and may ameliorate inflammation and liver enzyme levels and lead to a reduction in CVD mortality and morbidity, despite no proven benefit on liver histology [67].
It is well established that low-grade systemic inflammation, aside from contributing to CVD development, may also promote tumorigenesis. There is significant epidemiological evidence demonstrating that MASLD is a risk factor for the development of HCC, with current data indicating that HCC associated with MASLD may occur in a substantial number of patients who have not developed cirrhosis [70]. Even more concerning are findings that MASLD, as an independent factor, increases the risk of certain extrahepatic tumors. All these risks were found to be independent of factors such as age, sex, T2DM, obesity, and smoking. Results from a large meta-analysis by Mantovani et al. [71] demonstrated that the presence of MASLD is significantly associated with an increased risk of developing various cancers. This analysis, which encompassed 10 cohort studies with a total of 182,202 middle-aged subjects and approximately 8,500 cases of extrahepatic cancers at different sites over a median follow-up of 5.8 years, revealed that MASLD increases the risk of thyroid cancer by approximately 2.5-fold. The risk for developing extrahepatic gastrointestinal cancers, including cancers of the esophagus, stomach, pancreas, and colon, is increased by 1.5- to 2-fold. Moreover, MASLD has been shown to elevate the risk of tumors outside the digestive tract, with the incidence of breast, gynecological, lung, or urinary system cancers rising by 1.2- to 1.5-fold [71].
Early detection of MASLD and the identification of individuals at subsequent risk for developing various hepatic and extrahepatic complications highlight the importance of defining reliable biomarkers that could serve as early diagnostic tools, thereby enabling the timely implementation of both preventive and therapeutic strategies. All patients diagnosed with MASLD should be strongly encouraged to adopt primarily non-pharmacological interventions, with particular emphasis on promoting a healthy lifestyle, maintaining a balanced diet, avoidance of drinking and smoking, and engaging in regular physical activity in order to reduce the risk of disease progression and associated adverse outcomes, given that these factors contribute to inhibition of MASLD by diminishing oxidative stress and inflammation [36].
Integrating biomarkers into MASLD staging and monitoring: a framework
Incorporation of biomarkers into clinical algorithms for MASLD staging and treatment monitoring should follow a structured, evidence-based approach. Initial risk stratification begins with widely available non-invasive serum indices such as the fibrosis-4 (FIB-4) score and the NAFLD fibrosis score (NFS). Patients with indeterminate or elevated results should proceed to second-line testing, including elastography (transient or MRI-based) or serum panels such as the Enhanced Liver Fibrosis (ELF) test [72, 73]. For finer staging and detection of steatohepatitis, disease activity, or early fibrosis, markers such as cytokeratin-18 (CK-18), N-terminal propeptide of collagen type 3 (PRO-C3), and omics-based profiles (transcriptomic or lipidomic) can be used [74, 75]. Treatment response may be assessed using changes in imaging-based stiffness measures or serum biomarkers, including ELF dynamics or percentage reduction in magnetic resonance imaging-proton density fat fraction (MRI-PDFF) [72, 73]. Guidelines from the European Association for the Study of the Liver (EASL), the European Association for the Study of Diabetes (EASD), the European Association for the Study of Obesity (EASO), and regional hepatology associations recommend a stepwise algorithm: initial serum testing followed by liver ultrasound-based imaging techniques and fibrosis assessment [63, 76].
A proposal of integration of biomarkers into MASLD staging and treatment monitoring is presented in Figure 3.
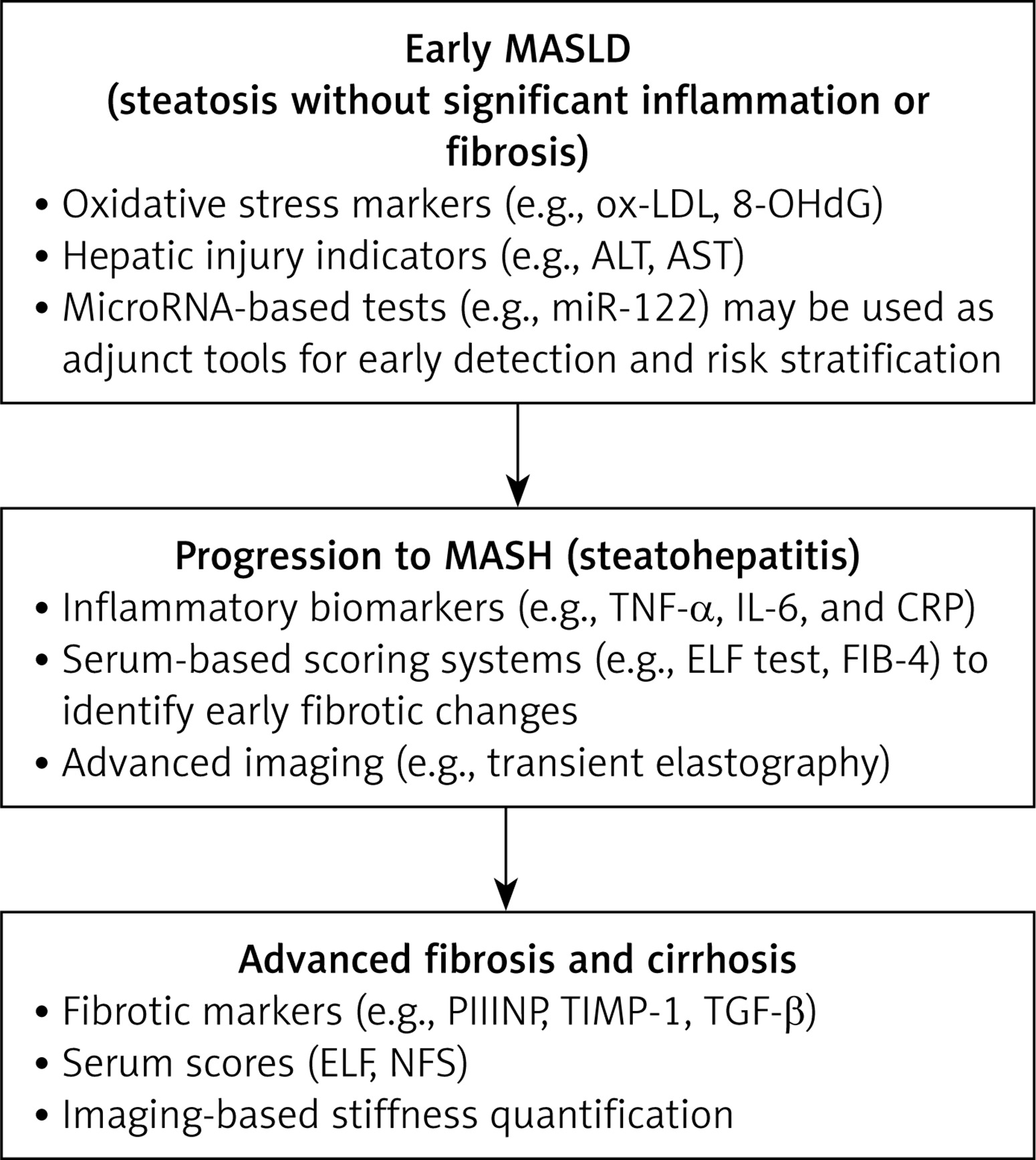
This figure illustrates the proposal of integration of biomarkers into MASLD staging and treatment monitoring (i.e., early MASLD, MASH, and advanced fibrosis and cirrhosis).
Clinical translation: biomarkers closest to bedside use in MASLD
As the field of MASLD biomarker research progresses, several candidates are emerging as practical tools for clinical translation [77]. Among non-invasive biomarkers, cytokeratin-18 (CK-18) fragments remain one of the most extensively studied indicators of hepatocyte apoptosis, demonstrating moderate diagnostic accuracy for differentiating MASH from simple steatosis [78]. Similarly, the ELF score, which combines serum markers of matrix turnover – including hyaluronic acid, procollagen type III n-terminal peptide (PIIINP), and tissue inhibitor of matrix metalloproteinases-1 (TIMP-1) – has been validated for fibrosis staging and is currently recommended by several guidelines [79, 80].
Serum biomarkers such as PRO-C3 (a marker of active fibrogenesis) are also gaining attention due to their specificity for collagen formation and their integration into composite indices such as the ADAPT score [81, 82]. In imaging, transient elastography (FibroScan) and magnetic resonance elastography (MRE) are increasingly accepted as reliable non-invasive tools for fibrosis assessment [83].
Integration of artificial intelligence in predictive modeling of MASLD progression
The integration of artificial intelligence (AI) and machine learning (ML) techniques into clinical hepatology has opened new possibilities for early detection and risk stratification of MASLD and its complications [84, 85]. Given the complexity and heterogeneity of MASLD pathogenesis, AI models are particularly suitable for identifying subtle, multidimensional patterns across clinical, biochemical, imaging, and multi-omics datasets [86–89].
Recent studies have demonstrated the utility of supervised ML algorithms in predicting key disease outcomes including progression from simple steatosis to MASH, development of significant fibrosis, and even HCC. These models often outperform traditional statistical methods in terms of sensitivity and specificity, especially when trained on large, diverse datasets that incorporate not only clinical and laboratory data but also transcriptomic, proteomic, and lipidomic signatures [90, 91].
Despite these advances, challenges remain, such as model interpretability, standardization across cohorts, and prospective validation. Nevertheless, AI-driven approaches represent a promising frontier in the management of MASLD, with the potential to enhance precision medicine strategies and enable timely intervention before irreversible liver damage occurs.
Ongoing trials and regulatory developments in MASLD
Significant advances have been made in the clinical development pipeline for MASLD, with several therapeutic agents progressing to late-stage trials and achieving key regulatory milestones. In March 2024, the U.S. FDA granted accelerated approval to resmetirom (Rezdiffra), a selective thyroid hormone receptor-β (THR-β) agonist, for the treatment of non-cirrhotic MASH. This approval represents a landmark achievement, marking the first FDA-approved pharmacological therapy specifically targeting MASH, based on evidence of fibrosis improvement without worsening of steatohepatitis [92, 93].
A number of agents – including semaglutide, resmetirom, survodutide, pegozafermin, denifanstat, obeticholic acid, tirzepatide, and the combination of cilofexor plus firsocostat – have demonstrated superiority over placebo in promoting fibrosis regression without exacerbating steatohepatitis. Notably, semaglutide, a glucagon-like peptide-1 (GLP-1) receptor agonist originally developed for T2DM and obesity, has shown robust efficacy in achieving resolution of steatohepatitis, underscoring its potential for repurposing in MASLD management [94]. Currently, a large number of ongoing clinical trials are being conducted worldwide to evaluate novel therapeutic strategies for MASLD, opening opportunities for the development of new drug candidates or combination therapies with synergistic effects or targeting distinct stages of disease progression. Some of these agents may emerge as effective treatment options. Despite the encouraging advances in therapeutic development, further research is urgently needed to establish more personalized and accessible treatment paradigms for MASLD in the near future [95].
Oxidative stress and inflammation biomarkers in MASLD
Oxidative stress and inflammation play pivotal roles in the pathogenesis of MASLD [96, 97]. This section aims to systematically examine oxidative stress and inflammation biomarkers in MASLD, highlighting current diagnostic advances and future perspectives in the field. By detailing the clinical applications of lipid peroxidation products, protein/DNA damage markers, and ROS, we seek to contribute to the development of non-invasive diagnostic approaches in MASLD. In addition to discussing the limitations of existing biomarkers, this section will evaluate the emerging potential of multi-omics approaches and innovative technologies in MASLD management.
Oxidative stress markers
Sources of reactive oxygen species (ROS) in MASLD
Multiple cellular sources contribute to excessive ROS generation in MASLD, acting synergistically to promote lipid peroxidation, inflammation, and hepatocellular damage. The major ROS-producing systems include mitochondrial electron transport chain dysfunction, cytochrome P450 2E1 (CYP2E1) activity, and NADPH oxidase (NOX) enzymes [98].
In the setting of insulin resistance and obesity, elevated FFA flux overwhelms hepatic mitochondrial β-oxidation capacity. This leads to electron leakage from complexes I and III of the electron transport chain (ETC), resulting in the formation of superoxide radicals (O2•–) [99]. The subsequent oxidative damage includes mitochondrial DNA (mtDNA) injury, opening of the permeability transition pore (mPTP), and initiation of apoptotic pathways [100]. Notably, 8-hydroxy-2′-deoxyguanosine (8-OHdG), a marker of oxidative DNA damage, is elevated in liver biopsy specimens of MASLD patients, and31P-MRS imaging has demonstrated ATP depletion consistent with mitochondrial dysfunction [101, 102]. While resmetirom remains the only FDA-approved therapy to date, experimental agents such as elamipretide (SS-31) and uncoupling protein 2 (UCP2) modulators are under investigation to restore mitochondrial integrity and reduce ROS production [103–106].
CYP2E1, an ROS-generating enzyme of the cytochrome P450 family, is overexpressed in MASLD and contributes to both mitochondrial and microsomal ROS accumulation. This creates a vicious cycle wherein oxidative stress further activates CYP2E1, aggravating lipid peroxidation, hepatocyte injury, and inflammation. Elevated serum CYP2E1 levels correlate with disease severity, and genetic polymorphisms such as the CYP2E1 c2 allele are associated with increased susceptibility to MASLD progression [107–109]. Promising therapeutic strategies include natural inhibitors (e.g., baicalin, silymarin, quercetin), pharmacological inhibitors like chlormethiazole, and gene-targeting approaches such as clustered regularly interspaced short palindromic repeat (CRISPR)-associated nuclease 9 (Cas9)-mediated knockdown in preclinical models [107, 110–112].
The NADPH oxidase (NOX) family also plays a central role in hepatic ROS generation [112]. Among its isoforms, NOX2 is predominantly expressed in Kupffer cells and is activated via toll-like receptor 4 (TLR4) signaling, resulting in superoxide production and pro-inflammatory cytokine release [113, 114]. In contrast, NOX4 is found in hepatocytes and hepatic stellate cells, where it promotes fibrogenesis through transforming growth factor-β (TGF-β) activation via hydrogen peroxide (H2O2) production. Circulating NOX4 levels have been shown to correlate with fibrosis stage in MASLD [113, 114]. Targeting NOX enzymes, especially NOX4, has shown promise in experimental models, and the dual NOX1/NOX4 inhibitor GKT137831 (setanaxib) has demonstrated anti-fibrotic efficacy in Phase II trials. Additional approaches, including siRNA-mediated NOX4 silencing and selective NOX1/4 inhibition, are being explored as potential therapeutic interventions [115, 116].
Together, these ROS sources not only drive liver injury but also serve as therapeutic targets for halting or reversing MASLD progression.
Table I and Figure 4 illustrate the sources of ROS in the pathogenesis of MASLD.
Table I
Sources of reactive oxygen species (ROS) in the pathogenesis of MASLD
| Sources of ROS | Mechanism | Clinical marker | Treatment candidates |
|---|---|---|---|
| Mitochondrial ETC [104] | Electron leakage → O2•– | mtDNA, 8-OHdG | Elamipretide, UCP2 agonists |
| CYP2E1 [107–109] | ω-hydroxylation → H2O2/4-HNE | Serum CYP2E1, MDA | Diallyl sulfide, chlormethiazole |
| NOX1/4 [115] | Cell type-specific ROS production | NOX4, TGF-β | GKT137831 |
[i] ROS – reactive oxygen species, O2•– – superoxide radical, mtDNA – mitochondrial DNA, 8OHdG – 8-hydroxy-2′-deoxyguanosine, UCP2 – uncoupling protein 2, CYP2E1 – cytochrome P450 2E1, NOX – NADPH oxidase, MDA – malondialdehyde, 4-HNE – 4-hydroxynonenal, TGF-β – transforming growth factor-β, H2O2 – hydrogen peroxide.
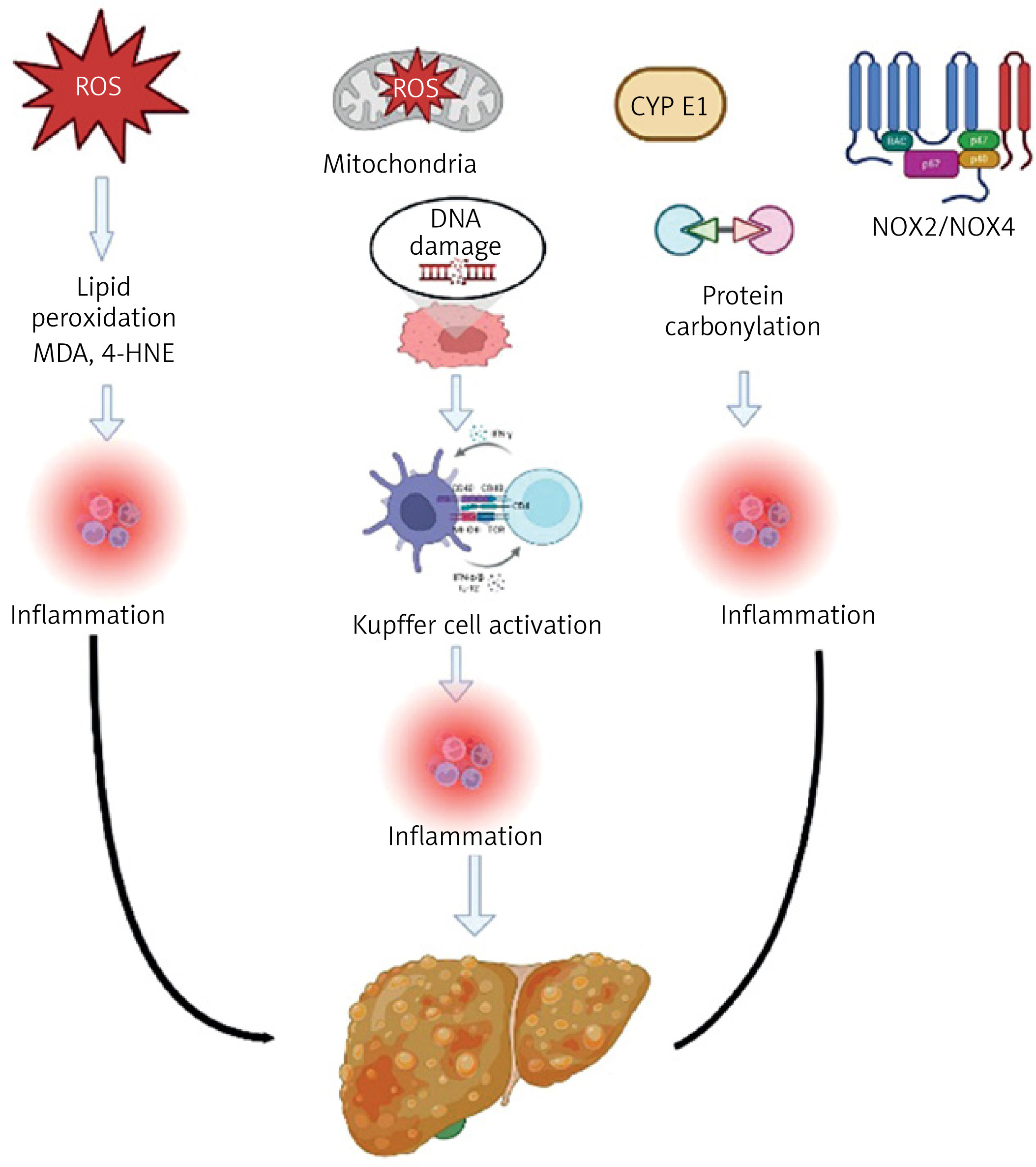
This illustration summarizes the key sources and consequences of ROS in MASLD. Major contributors to ROS production include mitochondrial dysfunction, cytochrome P450 2E1 (CYP2E1), and NADPH oxidase (NOX) enzymes. Excess ROS leads to lipid peroxidation, DNA and protein damage, and activation of Kupffer cells and hepatic stellate cells (HSCs). These events promote inflammation, apoptosis, and fibrogenesis, driving disease progression from simple steatosis to steatohepatitis (MASH) and ultimately cirrhosis.
Direct ROS measurement biomarkers
These biomarkers enable direct quantification of oxidative stress in MASLD; however, their application is limited by methodological complexities [117].
Hydroperoxides (ROOH)
Hydroperoxides are unstable compounds formed via lipid (LOOH), protein (POOH), and DNA oxidation, originating primarily from three hepatic sources: (1) electron leakage from the mitochondrial electron transport chain (particularly Complex I/III), (2) ROS-generating activity of CYP2E1, and (3) the NOX enzyme family (predominantly NOX4). Their short half-life and the requirement for immediate sample processing pose major limitations. Due to their instability, these compounds rapidly degrade into toxic end-products such as malondialdehyde (MDA) and 4-hydroxynonenal (4-HNE) [117, 118].
Future approaches may include nano-sensor technologies for real-time monitoring and stable isotope liquid chromatography-tandem mass spectrometry (LC-MS/MS) methods, which may enhance clinical utility and understanding of hydroperoxides in MASLD pathogenesis [119].
Hydroxyl radicals (•OH) and oxidative damage
Hydroxyl radicals (•OH) are the most reactive species among ROS and can cause severe oxidative damage in biological systems. These radicals are primarily generated through mechanisms such as the Fenton reaction and the Haber-Weiss cycle. In the Fenton reaction, hydroxyl radicals are produced via the reaction of ferrous ions (Fe2+) with hydrogen peroxide (H2O2). In the Haber-Weiss cycle, •OH is generated through the interaction of superoxide anion (O2•–) and H2O2. These processes contribute to increased oxidative stress in various tissues, including the liver [120].
Oxidative stress is known to play a significant role in the pathogenesis of MASLD. In patients with MASLD, decreased activity of antioxidant enzymes and increased ROS levels have been observed. This imbalance leads to an elevation in markers of oxidative damage such as lipid peroxidation, protein carbonylation, and DNA damage. Notably, DNA oxidation products such as 8-OHdG have been found at elevated levels in MASLD patients and have shown a correlation with disease severity [101, 121, 122].
Although direct measurement of hydroxyl radicals is challenging, oxidative damage can be assessed using indirect biomarkers. For example, levels of lipid peroxidation products and protein carbonylation can serve as indicators of oxidative stress. These biomarkers have been reported to increase with the progression of MASLD [3].
Innovative approaches such as enhancing the sensitivity of cryogenic ESR and the development of gadolinium-based MRI probes are being explored to better assess•OH-associated oxidative damage. These techniques may facilitate a deeper understanding of the pathological effects of•OH and improve its utility in clinical diagnostics [123].
Superoxide radical (O2•–) and oxidative damage
The superoxide radical (O2•–) is one of the primary precursors of ROS and is generated during normal cellular metabolism. Under physiological conditions, O2•– concentrations are approximately 0.1 nM; however, during inflammatory responses, these levels may increase to 100-fold, potentially resulting in damage to biomolecules [124, 125].
O2•– is detoxified by superoxide dismutase (SOD) enzymes, which convert it into H2O2. H2O2 is subsequently broken down into water and oxygen by the catalase (CAT) enzyme. These enzymatic defense mechanisms play a crucial role in maintaining the balance of ROS levels [124, 125].
Excessive production of O2•– and its derivatives contributes to damage in cellular components such as lipid peroxidation, protein carbonylation, and DNA oxidation, thereby triggering inflammation and promoting disease progression [126]. Therefore, biomarkers reflecting O2•– levels and the oxidative damage it induces may be valuable in the assessment of MASLD.
Hydrogen peroxide (H2 O2) and oxidative damage
Hydrogen peroxide (H2O2) is a key molecule among ROS, generated through various cellular sources such as mitochondrial respiration, peroxisomes, and cytochrome P450 enzymes. Through the Fenton reaction, H2O2 interacts with iron ions to produce highly reactive hydroxyl radicals (•OH), which can damage biomolecules including proteins, lipids, and DNA, thereby impairing cellular functions and leading to cell death [114].
As MASLD progresses, the excessive accumulation and metabolism of fatty acids lead to mitochondrial dysfunction, resulting in increased ROS production and oxidative stress. Oxidative stress triggers inflammatory responses, contributing to hepatocyte injury and disease progression [127].
Although direct measurement of H2O2 is challenging, oxidative stress can be assessed using indirect biomarkers. For example, advanced oxidation protein products (AOPP), total oxidant status (TOS), and oxidative-stress index (OSI) have been found to be elevated in MASLD patients and correlated with disease severity [128].
Clinical implications and future perspectives
Clinically, the assessment of ROS-related oxidative damage plays a critical role in predicting disease course and guiding therapeutic strategies. For example, elevated levels of O2•– and H2O2 are associated with systemic parameters such as TOS, total antioxidant capacity (TAC), and OSI. Accumulation of 8-OHdG, induced by hydroxyl radicals, serves as a marker of DNA damage and can be detected in both liver tissue and urine, correlating with disease activity. The major limitation in clinical use of these parameters is the short half-life and rapid degradation of the ROS molecules themselves. However, this limitation can be partially overcome through indirect measurements.
In the near future, sensitive and specific biomarkers for ROS-induced oxidative stress may offer new opportunities for early diagnosis and staging of MASLD. In this regard, technologies such as cryogenic electron spin resonance (ESR) may allow real-time in vivo monitoring of radical species. Stable isotope-labeled LC-MS/MS methods enable more accurate and specific quantification of H2O2 and lipid peroxidation products. Gadolinium-based magnetic resonance probes may permit imaging-based assessment of hepatic oxidative stress. Additionally, graphene-based electrochemical chips capable of simultaneously detecting O2•–, H2O2, and•OH, gas chromatography of exhaled breath (to detect volatile ROS metabolites), and PET tracers such as13F-ROStrace hold revolutionary potential for clinical application. These advances could mark the beginning of a new era in the early detection and monitoring of MASLD progression [129].
Lipid peroxidation products
Lipid peroxidation is one of the most critical outcomes of oxidative stress in the pathogenesis of MASLD. These products are formed through the interaction of ROS with lipids, leading to cell membrane damage, protein modification, and DNA injury, thereby accelerating disease progression [129].
Moreover, lipid peroxidation products may contribute to hepatic dysfunction and the development of MASLD through mechanisms involving inflammation and the dysregulation of insulin signaling. The accumulation of these products disrupts lipid metabolism and redox homeostasis, further promoting disease progression [129].
Malondialdehyde (MDA)
Malondialdehyde (MDA) is a final product of lipid peroxidation and is widely recognized as a key biomarker of oxidative stress [130]. MDA is generated through the reaction of ROS with polyunsaturated fatty acids in cellular membranes and is considered an indicator of cellular damage and inflammation [131].
In the pathogenesis of MASLD, the oxidation of accumulated fatty acids in hepatocytes leads to increased ROS production and subsequent formation of MDA [132]. This process triggers lipid peroxidation in hepatocytes, resulting in cellular injury and the initiation of inflammatory responses. MDA can form adducts by reacting with proteins and DNA, thereby disrupting cellular functions and potentially inducing mutations in genetic material [131].
Clinical studies have shown that MDA levels in serum and plasma samples of MASLD patients are significantly higher than in healthy individuals [133]. This increase correlates with disease severity, suggesting that MDA may play a role in the progression of MASLD. Notably, elevated MDA levels have also been observed in the early stages of the disease, indicating its potential utility as an early diagnostic biomarker for MASLD [134].
In MASLD patients, immune responses against lipid peroxidation products may serve as important indicators of progression to advanced stages such as fibrosis. For instance, high titers of IgG antibodies against MDA-modified human serum albumin (MDA-HSA) have been more prominently observed in patients with advanced fibrosis or cirrhosis [135]. This finding suggests that oxidative stress-induced immune responses may serve as independent markers of MASLD progression.
Numerous studies have indicated that MDA levels can be reduced through antioxidant therapies [136–140]. In the PIVENS trial, vitamin E (α-tocopherol) was found to reduce MDA levels and improve histological features in MASLD patients [141]. Polyphenols – such as resveratrol and extracts of green and white tea – have been demonstrated to inhibit lipid peroxidation in animal models [136–140].
Advanced oxidation protein products (AOPPs)
Advanced oxidation protein products (AOPPs) are compounds formed as a result of oxidative modifications of proteins and are considered important indicators of oxidative stress and inflammation. AOPPs are primarily generated through the action of chlorinated oxidants, particularly via the enzyme myeloperoxidase, and play a role in the pathogenesis of chronic inflammatory diseases [142].
Studies conducted in patients with MASLD have shown significantly elevated levels of AOPPs. This increase underscores the role of oxidative stress and inflammation in the progression of the disease. For instance, one study found that AOPP levels were markedly higher in individuals with MASLD and that this elevation was associated with cardiometabolic risk factors [128].
The measurement of AOPP levels is considered a potential tool for the early diagnosis of MASLD and for monitoring disease progression. Using AOPPs as markers of oxidative stress and inflammation may assist in evaluating disease advancement and response to treatment. However, further research is needed to clarify the specificity of AOPPs and their relationship with other biomarkers.
A better understanding of the role of AOPPs in MASLD pathogenesis could contribute to the development of new therapeutic strategies. It has been suggested that antioxidant therapies may alleviate oxidative stress by reducing AOPP levels, thereby slowing disease progression. For example, antioxidants such as vitamin E have been reported to stabilize AOPP levels, although they do not appear to significantly affect the progression of atherosclerosis [143].
4-Hydroxynonenal (4-HNE)
4-Hydroxynonenal (4-HNE) is a major end product of lipid peroxidation and is considered a biomarker of oxidative stress. It is formed through the reaction of ROS with polyunsaturated fatty acids in cellular membranes. 4-HNE can form covalent bonds with proteins, DNA, and phospholipids, thereby disrupting cellular functions and playing a role in the pathogenesis of various diseases [144].
The use of 4-HNE as an indicator of oxidative stress and lipid peroxidation is particularly important in diseases such as MASLD. The accumulation of 4-HNE-protein adducts in tissues reflects the degree of oxidative damage and may provide insight into the progression of the disease [145].
Measurement of 4-HNE levels is considered a potential tool for the early diagnosis and monitoring of MASLD. However, due to its high reactivity and short half-life, direct measurement of 4-HNE is challenging. Therefore, indirect measurements such as the detection of 4-HNE-protein adducts via immunohistochemical methods or specific assays like ELISA are preferred [146].
A better understanding of the role of 4-HNE in the pathogenesis of MASLD may contribute to the development of new therapeutic strategies. In particular, therapeutic approaches aimed at enhancing the activity of enzymes involved in the detoxification of 4-HNE – such as aldehyde dehydrogenase (ALDH) and glutathione (GSH) – may offer potential benefits in reducing oxidative stress. For example, the use of ALDH2 activators may accelerate the detoxification of 4-HNE and reduce cellular damage [130, 147].
F2-Isoprostanes
F2-Isoprostanes are prostaglandin-like compounds formed by the non-enzymatic peroxidation of arachidonic acid by free radicals and are considered reliable biomarkers of in vivo lipid peroxidation [148]. Their stable structure and specific formation mechanisms make F2-isoprostanes sensitive and reliable indicators of oxidative stress. Levels of these compounds are elevated in various diseases associated with oxidative stress, highlighting their significance as biomarkers.
Measurement of F2-isoprostane levels in MASLD patients can provide valuable information about the severity and progression of the disease. Specifically, elevated urinary 8-iso-PGF2α levels have been correlated with the degree of liver steatosis. This finding suggests that F2-isoprostanes may be a potential tool for the non-invasive assessment of MASLD [149].
A better understanding of the role of F2-isoprostanes in the pathogenesis of MASLD may contribute to the development of new therapeutic strategies. In particular, antioxidant treatments aimed at reducing oxidative stress may slow disease progression by lowering F2-isoprostane levels. For example, omega-3 fatty acid supplementation has been shown to reduce F2-isoprostane levels and alleviate oxidative stress [150].
Oxidized LDL (ox-LDL)
Oxidized low-density lipoprotein (ox-LDL), generated through the oxidative modification of LDL particles by ROS and metal ions, is a key product of lipid peroxidation that plays an active role in the pathogenesis of MASLD. Unlike free lipid peroxidation products such as MDA or 4-HNE, ox-LDL circulates systemically and engages in pathogenic signaling by activating Kupffer cells, promoting NLRP3 inflammasome activation, and inducing proinflammatory cytokines like TNF-α and IL-1β [151, 152].
These effects contribute to hepatic inflammation, oxidative damage, and fibrogenesis. Elevated circulating ox-LDL levels have been associated with early atherogenesis and may also reflect hepatic oxidative stress in MASLD, making ox-LDL a promising non-invasive biomarker for both hepatic and cardiovascular risk stratification in this patient population [153, 154].
From a therapeutic standpoint, statins, proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitors, and antioxidants have shown potential to reduce ox-LDL levels or mitigate its harmful effects. Furthermore, targeting ox-LDL or its receptors (e.g., LOX-1) is being explored as a novel strategy to interrupt the oxidative-inflammatory cascade in MASLD [136, 152].
Antioxidant depletion markers
In patients with MASLD, significant reductions in the levels of various antioxidants have been observed [155]. Specifically, decreases in the activities of enzymatic antioxidants such as glutathione (GSH), SOD, CAT, and glutathione peroxidase (GPx) have been reported [156, 157]. A decrease in these enzymes can increase oxidative stress and exacerbate cellular damage.
Measurement of antioxidant levels is considered a potential tool for the early diagnosis and monitoring of disease progression in MASLD [158]. For example, plasma coenzyme Q10 and SOD levels have been found to negatively correlate with necroinflammatory activity and the degree of fibrosis [159]. This suggests that antioxidant markers may assist in evaluating the severity and prognosis of the disease.
In the treatment of MASLD, strategies aimed at strengthening antioxidant defense mechanisms show promise. Research is ongoing on agents that enhance the activity of antioxidant enzymes or elevate the levels of antioxidant molecules. Furthermore, the positive effects of dietary antioxidants, such as caffeic acid and vitamin E, on MASLD are being investigated [141, 160].
Glutathione (GSH) and oxidized glutathione (GSSG)
Oxidative stress plays a central role in the pathogenesis of MASLD, and GSH functions as an important antioxidant in this process [130]. GSH is a key component of cellular antioxidant defense. In its γ-glutamyl-cysteinyl-glycine structure, GSH neutralizes ROS, such as H2O2 and lipid peroxides, through its sulfhydryl (-SH) group [130]. It works in conjunction with enzymes such as GPx and glutathione-S-transferase (GST). GSH plays a critical role in maintaining cellular redox balance and is converted to its oxidized form, GSSG, under oxidative stress conditions. The GSH/GSSG ratio is considered an indicator of cellular redox status; in healthy cells, this ratio is approximately 100 : 1, but it can decrease to 10 : 1 or 1 : 1 under oxidative stress [161, 162]. In MASLD patients, hepatic GSH levels are decreased, and the GSH/GSSG ratio is reduced [161]. These changes indicate increased oxidative stress and insufficient antioxidant defense mechanisms. Therefore, GSH levels and the GSH/GSSG ratio are considered potential biomarkers for the early diagnosis and monitoring of disease progression in MASLD. Specifically, a decrease in GSH levels and a drop in the GSH/GSSG ratio may indicate disease progression. In liver biopsy, GSH levels show an inverse correlation with the severity of MASH [97]. Additionally, a negative relationship has been found between the GSH/GSSG ratio and insulin resistance and fibrosis stage [161].
In MASLD treatment, strategies aimed at increasing GSH levels show promise. The use of antioxidant compounds, such as N-acetylcysteine (a precursor of GSH), to increase GSH levels may reduce oxidative stress and inflammation, positively affecting disease progression. Moreover, direct GSH supplementation has been shown to improve liver functions and reduce oxidative stress markers [163].
Superoxide dismutase (SOD) and catalase (CAT)
Oxidative stress plays a significant role in the pathogenesis of MASLD, and antioxidant enzymes such as SOD and CAT are crucial in this process. These enzymes reduce cellular oxidative stress by detoxifying superoxide anions and H2O2 [164]. An experimental study showed both biochemically and histopathologically that CAT deficiency increases lipid accumulation, inflammation, and fibrosis in the liver [165]. Similarly, in another experimental study, increased lipid accumulation in the liver was observed in SOD2 knockout mice [166].
SOD and CAT are key antioxidant enzymes that mitigate oxidative stress by converting superoxide anions into H2O2 and subsequently decomposing H2O2 into water and oxygen. In MASLD, alterations in their activity have been inconsistently reported. Some studies, particularly in early disease stages or experimental models, indicate upregulation of SOD and CAT as a compensatory response to rising ROS [167]. Conversely, in advanced stages such as fibrotic NASH, prolonged oxidative burden may deplete these enzymes, resulting in reduced activity. This biphasic pattern suggests that their diagnostic and prognostic utility depends on disease stage and timing of assessment. Therefore, while SOD and CAT are considered promising oxidative stress biomarkers for MASLD, standardized protocols and longitudinal studies are needed to clarify their clinical relevance [168, 169].
In MASLD treatment, strategies aimed at reducing oxidative stress are gaining importance. Pharmacological agents or genetic approaches that modulate the activities of SOD and CAT enzymes are being considered as potential therapeutic targets. Additionally, the effects of diet and lifestyle changes, particularly nutritional regimens that increase antioxidant capacity, on SOD and CAT activities are being investigated. For example, the Dietary Approaches to Stop Hypertension (DASH) diet has been shown to increase TAC and SOD levels in MASLD patients [170].
Vitamin E (α-tocopherol)
Vitamin E, known in its α-tocopherol form, is a potent lipophilic antioxidant. It plays a critical role in reducing oxidative stress by protecting cell membranes from free radical-induced peroxidative damage. The antioxidant effect of vitamin E stabilizes cell membranes and prevents damage caused by free radicals [171]. In MASLD, hepatic α-tocopherol levels decrease, and in experimental models, vitamin E supplementation has reduced inflammation and hepatic fibrosis [169]. Some clinical studies have shown that vitamin E provides histological improvement in MASH [141].
Although dietary antioxidant supplementation has been explored as a potential therapeutic strategy in MASLD, several limitations restrict its clinical applicability. First, the bioavailability and hepatic delivery of many dietary antioxidants (e.g., polyphenols, vitamins C and E) are often low and highly variable between individuals. Second, the antioxidant effects observed in vitro or in animal models may not translate to meaningful clinical outcomes in humans due to complex metabolic interactions. Moreover, supplementation studies frequently lack standardized dosages, durations, or endpoints, and some antioxidants may even exert pro-oxidant effects at high doses. Thus, while dietary antioxidants may offer supportive benefits, they should not be considered reliable surrogates for therapeutic efficacy without robust clinical trial evidence.
Coenzyme Q10 (ubiquinone)
Coenzyme Q10 (CoQ10) is a compound that plays an important role in mitochondrial energy production and is also a powerful antioxidant. With age, its endogenous production decreases, and it is used as a supportive treatment for conditions such as cardiovascular diseases, neurodegenerative diseases, and diabetes. In a clinical study, 240 mg/day CoQ10 treatment for 6 months significantly improved liver steatosis in MASLD patients [172]. However, the results of a meta-analysis including patients with metabolic syndrome who received CoQ10 supplementation showed significant improvements in liver enzymes, adiponectin levels, pro-inflammatory cytokine levels, and glucose levels, but the effect on MASLD remained limited [173].
Thioredoxin system
Thioredoxin is a protein system that regulates cellular redox balance and reduces oxidative stress. The thioredoxin system controls intracellular redox status, ensuring proper protein folding and maintaining their functions. Dysfunction of this system can contribute to increased oxidative stress and the progression of MASLD [174].
DNA/protein damage markers
Oxidative stress-induced DNA and protein damage in MASLD are important indicators of disease progression [130].
8-Hydroxy-2′-deoxyguanosine (8-OHdG)
8-OHdG is a common marker of oxidative DNA damage. It results from the effect of reactive oxygen species on the guanosine base in DNA and is typically detected in urine. Elevated levels of 8-OHdG may indicate oxidative stress and, consequently, the progression of MASLD [101, 175].
Measurement of 8-OHdG levels is used to assess oxidative stress and DNA damage. It can be detected in urine, serum, or tissue samples and has been associated with various cancers, neurodegenerative diseases, and inflammatory conditions [176]. Particularly, in urine samples, it can be measured with high sensitivity using methods such as LC-MS/MS [122]. Non-invasive measurement of 8-OHdG presents potential for early diagnosis of MASLD and monitoring treatment response. Moreover, therapeutic strategies aimed at reducing oxidative stress may contribute to disease management by lowering 8-OHdG levels [162, 163].
Protein carbonyls
Protein carbonyls contain ketone and aldehyde groups formed due to oxidation of the side chains of proteins, and they are an indicator of oxidative protein damage. These modifications can lead to loss of protein function and cellular dysfunction [177].
Protein carbonyls are used as biomarkers to assess oxidative stress and protein damage. They can be detected in serum or plasma samples and have been associated with aging, diabetes, Alzheimer’s disease, neonatal brain injury, and chronic inflammatory conditions [177, 178]. Monitoring protein carbonyl levels can be useful in understanding the pathogenesis of MASLD and evaluating the effectiveness of treatment strategies. Additionally, therapeutic approaches aimed at reducing protein carbonylation may positively influence the progression of the disease [177].
To provide a comprehensive yet structured overview of key oxidative stress-related biomarkers in MASLD, Tables II and III summarize their mechanistic roles, therapeutic relevance, biological sources, detection methods, clinical utility, and limitations. While these biomarkers hold diagnostic promise, a major challenge limiting the clinical adoption of oxidative stress biomarkers in MASLD is the lack of assay standardization. For instance, ELISA kits for 8-OHdG vary considerably in terms of antibody specificity and detection thresholds, leading to inter-laboratory variability. Similarly, although LC-MS/MS offers high sensitivity and specificity for markers such as F2-isoprostanes or 4-HNE, its widespread use is hampered by cost, technical complexity, and the need for highly trained personnel. The absence of universally accepted reference ranges and calibration standards further complicates cross-study comparisons and limits the translation of these biomarkers into routine clinical practice.
Table II
Summary of oxidative stress-related biomarkers and therapeutic targets in MASLD
| Biomarker/Target | Pathophysiological role | Therapeutic implications | Key references |
|---|---|---|---|
| MDA (malondialdehyde) | End-product of lipid peroxidation; reflects oxidative membrane damage | Antioxidants (e.g., vitamin E) reduce levels | [133, 134] |
| 4-HNE (4-hydroxynonenal) | Highly reactive aldehyde; modifies proteins/DNA; promotes inflammation | ALDH2 activators promote detoxification | [144–147] |
| F2-Isoprostanes | Stable lipid peroxidation marker; correlates with disease severity | Reduced by omega-3 fatty acids; clinical monitoring possible | [149, 150] |
| 8-OHdG (8-hydroxy-2′-deoxyguanosine) | DNA oxidation marker; correlates with fibrosis and disease activity | Potential non-invasive marker; lowered by antioxidant therapy | [101, 122] |
| Oxidized LDL (ox-LDL) | Links oxidative stress to systemic inflammation and CVD risk | Targeted by statins, PCSK9 inhibitors, anti-oxLDL immunotherapies | [152–154] |
| CYP2E1 | Microsomal and mitochondrial ROS production; promotes lipid peroxidation and apoptosis | Inhibitors: chlormethiazole, natural compounds, CRISPR strategies | [107–110] |
| NOX enzymes (NOX2/NOX4) | Promotes superoxide/H2O2 production; activates Kupffer cells and stellate cells | Setanaxib (NOX1/4 inhibitor), siRNA therapies | [113–116] |
| Mitochondrial ROS | ETC leakage causes O2•–; triggers apoptosis via mitochondrial damage | Targeted by elamipretide (SS-31), UCP2 modulation | [99–106] |
| GSH/GSSG ratio | Indicates antioxidant depletion; inversely correlates with disease severity | N-acetylcysteine and other GSH precursors may restore balance | [161–163] |
| SOD / CAT | Antioxidant enzymes defending against ROS; imbalance promotes damage | Modulated by diet/lifestyle; DASH increases SOD | [167, 168, 170] |
| Vitamin E, CoQ10 | Supplemental antioxidants: reduce ROS and improve liver markers | Used in adjunct therapy; mixed clinical results | [136, 141, 172, 173] |
[i] MASLD – metabolic dysfunction-associated steatotic liver disease, ROS – reactive oxygen species, MDA – malondialdehyde, 4-HNE – 4-hydroxynonenal, 8-OHdG – 8-hydroxy-2’-deoxyguanosine, ox-LDL – oxidized low-density lipoprotein, NOX – NADPH oxidase, CYP2E1 – cytochrome P450 2E1, TGF-β – transforming growth factor β, UCP2 – uncoupling protein 2.
Table III
Key oxidative stress markers in MASLD: sources, detection, and clinical relevance
| Oxidative marker | Primary source | Detection methods | Clinical utility | Limitations | Key references |
|---|---|---|---|---|---|
| MDA | Lipid peroxidation of polyunsaturated fatty acids | TBARS assay, HPLC, ELISA | Widely used marker of lipid peroxidation; correlates with disease severity | Non-specific; affected by diet and sample handling | [133, 134] |
| 4-HNE | Degradation of ω-6 polyunsaturated fatty acids | ELISA, GC-MS, immunohistochemistry | Early indicator of oxidative injury and apoptosis | Short half-life; requires stabilization for accurate quantification | [144–147] |
| 8-OHdG | Oxidative DNA damage (nuclear and mtDNA) | ELISA, LC-MS/MS, immunostaining | Biomarker for DNA oxidative stress and fibrosis progression | May lack tissue specificity; influenced by comorbidities | [101, 122] |
| Oxidized LDL (ox-LDL) | Oxidatively modified circulating LDL particles | ELISA, chemiluminescence, flow cytometry | Reflects both oxidative stress and systemic inflammation; CVD linkage | Assay variability; lack of standardization in clinical practice | [152–154] |
| F2-Isoprostanes | Non-enzymatic oxidation of arachidonic acid | GC-MS, LC-MS, ELISA | Sensitive marker of oxidative damage; useful in early detection | Technical complexity; limited availability in clinical labs | [149, 150] |
[i] MASLD – metabolic dysfunction-associated steatotic liver disease, MDA – malondialdehyde, 4-HNE – 4-hydroxynonenal, 8-OHdG – 8-hydroxy-2′-deoxyguanosine, ox-LDL – oxidized low-density lipoprotein, F2-isoprostanes – F2 class of prostaglandin-like compounds formed by lipid peroxidation, ELISA – enzyme-linked immunosorbent assay, HPLC – high-performance liquid chromatography, GC-MS – gas chromatography-mass spectrometry, LC-MS/MS – liquid chromatography-tandem mass spectrometry.
Inflammatory markers in MASLD
Inflammatory pathways in MASLD
Inflammation plays a critical role in the progression of MASLD. The inflammatory responses in MASLD are typically triggered through various cellular pathways. Some of these pathways include the activation of Kupffer cells, dysfunction of adipose tissue, and inflammasome activation (especially the NLRP3 inflammasome). To provide a structured comparison of the inflammatory biomarkers discussed in MASLD, Table IV summarizes their biological roles, diagnostic relevance, and current limitations based on available evidence.
Table IV
Comparative overview of inflammatory biomarkers in MASLD
| Biomarker | Role in MASLD pathogenesis | Diagnostic value | Limitations | Key references |
|---|---|---|---|---|
| TNF-α | Promotes hepatocyte apoptosis and insulin resistance | Correlates with disease severity; elevated in NASH | Lacks specificity; elevated in other inflammatory states | [96, 190] |
| IL-6 | Induces hepatic inflammation; associated with steatosis severity | Useful in differentiating MASLD stages | Affected by obesity, infection, and metabolic syndrome | [96, 193–195] |
| IL-1β | Enhances inflammasome activation and liver injury | Associated with necroinflammatory activity | Low stability; assay variability | [96] |
| CRP | Acute-phase protein; reflects systemic inflammation | Non-specific but elevated in MASLD and NASH | Elevated in many chronic diseases | [199, 201] |
| CCL2 (MCP-1) | Recruits monocytes to liver; correlates with steatohepatitis | Strong marker for NASH and fibrosis prediction | Assay standardization issues | [196, 197] |
[i] MASLD – metabolic dysfunction-associated steatotic liver disease, TNF-α – tumor necrosis factor-α, IL – interleukin, CRP – C-reactive protein, CCL2 – C-C motif chemokine ligand 2, MCP-1 – monocyte chemoattractant protein-1, CXCL10 – C-X-C motif chemokine ligand 10, ICAM-1 – intercellular adhesion molecule 1.
Kupffer cell activation
Kupffer cells are the resident macrophages of the liver and regulate the inflammatory response within the liver. In MASLD, fat accumulation and oxidative stress in the liver lead to the activation of Kupffer cells. Activated Kupffer cells secrete pro-inflammatory cytokines such TNF-α, interleukin (IL)-6, and IL-1β. These cytokines cause damage to liver cells and contribute to the spread of inflammation. Furthermore, Kupffer cells increase oxidative stress and inflammation, thus contributing to the progression of the disease [179].
Adipose tissue dysfunction
Adipose tissue is an important endocrine organ responsible for storing fat and maintaining energy balance in the body [180, 181]. A strong relationship exists between obesity and MASLD, as excessive fat accumulation leads to adipose tissue dysfunction [182]. Adipokines secreted by dysfunctional adipose tissue, particularly molecules such as leptin and adiponectin, increase inflammation [183]. Elevated leptin levels promote inflammation, while decreased adiponectin levels reduce anti-inflammatory effects. These changes pave the way for the progression of steatosis and the development of hepatitis in the liver [184].
NLRP3 inflammasome activation
The NLRP3 inflammasome is a protein complex that is activated in response to intracellular pathogens and stresses. In MASLD, oxidative stress, particularly associated with hepatic steatosis, triggers the activation of the NLRP3 inflammasome [185]. When the NLRP3 inflammasome is activated, it activates caspase-1, which then forms active forms of pro-inflammatory cytokines such as IL-1β and IL-18 [186]. These cytokines increase inflammation in the liver, leading to steatohepatitis and fibrosis. Moreover, inhibition of the NLRP3 inflammasome has emerged as a potential approach in the treatment of MASLD [187, 188].
Important inflammatory markers
Cytokines and chemokines
Various inflammatory markers play a crucial role in the pathogenesis of MASLD. Below are explanations of three inflammatory markers that are critical in MASLD: TNF-α, IL-6, and chemokine (C-C motif) ligand 2 (CCL2) (also referred as monocyte chemoattractant protein-1, MCP-1).
Tumor necrosis factor-α (TNF-α): TNF-α is an important pro-inflammatory cytokine that plays a critical role in the development and progression of MASLD. One of the inflammatory mediators released during liver damage, TNF-α participates in the inflammatory cascade, triggering processes such as hepatocyte dysfunction, necrosis, and apoptosis. This can lead to the development of progressive diseases such as liver fibrosis and cirrhosis. Specifically, the induction of TNF-α by bacterial endotoxins is a critical factor in the progression of liver damage. In the context of MASLD, TNF-α works together with other cytokines such as IL-1 and IL-6 to maintain and amplify the liver’s inflammatory response. This inflammatory environment is a central factor contributing to disease progression and liver cell damage. Additionally, TNF-α plays an important role in the development of insulin resistance, which occurs when the body’s cells cannot respond to insulin, and the increase in inflammatory cytokines such as TNF-α contributes to this metabolic dysfunction. Elevated TNF-α levels are associated with both systemic inflammation and metabolic disorders specific to MASLD. Excessive expression of TNF-α triggers hepatocyte apoptosis, necrosis, and increased liver damage in animal models, indicating that this cytokine is not only an inflammation marker but also plays a direct role in the progression of liver disease, laying the groundwork for more severe conditions [189]. TNF-α can be considered an “alarm signal” that activates immune cells and other molecules during liver stress. However, when this signal is too strong or prolonged, it damages liver cells, leading to disease progression. This mechanism contributes to the progression of liver diseases such as MASLD by promoting liver cell death (apoptosis) and necrosis [96]. In animal models, inhibition of TNF-α has been shown to reduce liver steatosis and inflammation [190]. Anti-TNF-α therapies have shown promise in experimental MASLD models, where agents such as infliximab significantly reduced hepatic inflammation, fibrosis, and serum transaminase levels in rodent models of steatohepatitis [189]. These effects were associated with decreased hepatic TNF-α, IL-6, and IL-1β levels and improved insulin signaling. Although clinical data are limited, case observations – such as biochemical improvement in a MASH patient treated with adalimumab for comorbid rheumatoid arthritis – suggest potential translational relevance [191]. Furthermore, a recent study investigating patients with psoriatic arthritis (PsA) demonstrated that various biologic agents – including infliximab, adalimumab, etanercept, golimumab, certolizumab, ustekinumab, and secukinumab – were associated with an overall protective effect against liver fibrosis in individuals with MASLD. This study also highlighted the potential utility of the fibrosis-4 (FIB-4) score as a noninvasive screening tool for liver injury in this population [192]. However, the clinical use of direct TNF-α inhibitors for MASH has not yet been approved. While targeting TNF-α remains an attractive idea for reducing liver inflammation and insulin resistance in MASLD, more research and clinical studies are needed to create safe and effective interventions that can be translated into routine clinical practice.
Interleukin-6 (IL-6): IL-6 is a cytokine with both pro-inflammatory and anti-inflammatory properties, playing a crucial role in immune responses. In MASLD patients, IL-6 levels are typically elevated, which is associated with increased IL-6 production in the liver compared to healthy individuals [96]. High IL-6 levels contribute to liver inflammation, which, if persistent, can lead to liver cell damage and fibrosis [193]. IL-6 also plays a role in the recruitment of immune cells to the liver, which, while an important defense mechanism, can lead to excessive inflammation and further liver damage. Moreover, IL-6 is known to cause insulin resistance, which exacerbates MASLD [194].
Studies in animal models have shown that the absence or inhibition of IL-6 signaling reduces liver inflammation but does not completely prevent fat accumulation in liver cells [195]. These findings suggest that IL-6 plays a significant inflammatory role in liver diseases but is not sufficient on its own to prevent fat accumulation. In the absence of IL-6, reduced liver inflammation and less damage have been observed, but fat accumulation in the liver is not entirely halted. These findings highlight the important role of IL-6 in regulating inflammatory processes while also emphasizing that other factors, such as fat accumulation, must still be considered. Targeting IL-6 may help control some harmful processes in MASLD, but it also underscores the need for a broader approach in the treatment of the disease.
C-C motif chemokine ligand 2 (CCL2): C-C motif chemokine ligand 2 (CCL2), also known as monocyte chemoattractant protein-1 (MCP-1), is a chemokine that attracts monocytes to sites of inflammation. In MASLD, elevated CCL2 levels exacerbate liver inflammation by enhancing monocyte and macrophage infiltration [196]. Additionally, TNF-α has been shown to induce CCL2 production, and this mechanism is considered significant in metastatic liver diseases [197].
Acute phase proteins
Acute phase proteins are proteins whose levels rapidly increase in response to inflammation or tissue injury and play a role in modulating the immune response. These proteins are considered important biomarkers in the management of diseases such as MASLD. In MASLD, the levels of acute phase proteins are used to assess the presence and extent of inflammation, providing insight into the severity and progression of the disease.
C-reactive protein (CRP): C-reactive protein (CRP) is one of the most common and well-known acute-phase proteins [198]. It is rapidly produced by the liver in response to inflammation through IL-6 signaling and increases in response to bacterial infections, injuries, or tissue damage [199, 200]. In liver diseases such as MASLD, elevated CRP levels indicate an ongoing inflammatory process in the body. CRP can be considered a biomarker, particularly in the early stages of MASLD, such as hepatic steatosis and inflammation [199]. High CRP levels are associated with disease progression and are used as an indicator of liver inflammation [201]. By lowering CRP levels, CRP apheresis may help mitigate the adverse effects of elevated CRP concentrations on organs, including the liver. However, further studies are needed to determine whether this therapeutic approach would be effective in the treatment of chronic conditions such as MASLD [199].
Ferritin: Ferritin is a protein responsible for iron storage in the body and is produced by the liver as part of the acute-phase response [202]. Ferritin levels increase in inflammatory conditions. Hepatic iron accumulation leads to the production of ROS via the Fenton reaction, ultimately causing ferroptosis. It also exerts a pro-inflammatory effect by promoting IL-1β secretion from macrophages [203]. Elevated ferritin levels in MASLD may indicate hepatic inflammation and cellular injury [132]. Ferritin levels have been associated with an increased risk of fibrosis [204]. In addition, elevated ferritin may reflect hepatic steatosis and excessive iron accumulation. Ferritin can affect hepatic metabolic function, particularly in the context of inflammation and oxidative stress [184]. However, elevated ferritin reflects the complex interplay between iron metabolism and inflammation, making it a potentially valuable biomarker for monitoring the course of MASLD. Nevertheless, its specificity is limited in MASLD due to increased levels also being observed in conditions such as hemochromatosis/HFE mutations, alcohol-related liver disease, and infections. Therefore, ferritin may be more useful when interpreted alongside other acute-phase markers such as CRP during initial screening assessments. Regular monitoring of ferritin levels may also provide additional benefits in evaluating progression to cirrhosis and HCC and in determining the need for liver biopsy [205].
Fibrinogen: Fibrinogen is another acute-phase protein produced by the liver that plays a key role in blood coagulation. Its levels increase during inflammation and initiate coagulation mechanisms when vascular injury occurs. High levels of fibrinogen are likely to cause thrombosis. They also may activate platelets. In MASLD, elevated fibrinogen levels indicate the liver’s response to inflammation and fibrosis. This increase may also reflect a heightened risk for additional complications, such as bleeding and coagulation disorders, associated with hepatic inflammation [206]. Furthermore, fibrinogen levels may serve as an early indicator of the transition to fibrotic stages of MASLD. Its hepatic production is upregulated via the IL-6/STAT3 signaling pathway, and it facilitates fibrosis by enhancing hepatic stellate cell activation [207]. Fibrinogen levels exceeding 400 mg/dl have been associated with portal hypertension [208].
Serum amyloid A (SAA): Serum amyloid A (SAA) is a protein that rapidly increases during the early stages of inflammation and is primarily produced by the liver. As part of the acute-phase protein response, SAA participates in the body’s inflammatory reaction and is commonly used to monitor the course of various inflammatory diseases. In MASLD, elevated SAA levels may reflect hepatic inflammation and the resulting tissue damage associated with this inflammatory process [209]. SAA can be considered an important biomarker, particularly in the more advanced stages of MASLD, such as MASH. High SAA levels indicate ongoing inflammation and suggest that disease progression should be closely monitored [210].
Adipokines
Adipokines are biologically active molecules secreted by adipose (fat) tissue that regulate energy balance, metabolism, inflammation, and immune responses in the body. These molecules are diverse, including leptin, adiponectin, and resistin, and can generally exhibit either pro-inflammatory or anti-inflammatory properties. The dysfunction of adipokines can contribute to the development of diseases such as obesity and MASLD [183].
Leptin: Leptin, primarily secreted by adipocytes, plays a key role in regulating energy balance, appetite, and metabolic processes. It is often referred to as the “satiety hormone” because it helps to suppress hunger and increase energy expenditure. However, in the context of metabolic dysfunction, including conditions such as obesity and MASLD, leptin’s role becomes more complex [211].
In MASLD, leptin levels are often elevated, particularly in obese individuals [184]. This increase in leptin is generally a result of the larger adipose tissue mass commonly found in individuals with metabolic dysfunction. While leptin’s role in regulating appetite and energy expenditure is well known, its involvement in inflammation and IR has gained increasing attention in recent years [212].
Leptin is known to have pro-inflammatory properties. It activates several inflammatory pathways, including the nuclear factor kappa B (NF-κB) pathway, which is involved in immune cell activation and the release of inflammatory cytokines. In MASLD, elevated leptin levels contribute to the development of IR, a hallmark feature of the disease. Leptin interferes with the insulin signaling pathway, leading to impaired glucose and lipid metabolism, and promotes liver fat accumulation, which is characteristic of MASLD [213, 214].
Moreover, leptin promotes the activation of Kupffer cells, the liver’s resident macrophages, enhancing the release of inflammatory cytokines such as TNF-α, IL-6, and IL-1β. This creates a vicious cycle where inflammation and IR perpetuate the progression of MASLD. Additionally, leptin is also associated with hepatic steatosis and fibrosis. Elevated leptin levels in the circulation are correlated with the severity of liver damage, including progression from simple steatosis to MASH and even cirrhosis in some cases [184].
Interestingly, despite leptin’s role in promoting inflammation and IR, it also exhibits some beneficial effects in energy-deficient conditions. Under normal circumstances, leptin signals to the brain to reduce appetite and increase energy expenditure. However, in the setting of MASLD, where leptin resistance is often present, the body no longer responds effectively to leptin’s signals. This can lead to overeating and further exacerbate the metabolic disturbances contributing to the disease [215].
In summary, leptin acts as an important mediator in the pathogenesis of MASLD, serving as both a marker of obesity and a regulator of inflammation and IR. Elevated leptin levels observed in MASLD patients are associated with worsened liver damage, and targeting leptin signaling pathways may offer a potential therapeutic strategy for managing MASLD. Further studies are needed to better understand leptin’s complex role in MASLD and its potential as a therapeutic target for preventing or treating the disease [215].
Adiponectin: Adiponectin is a protective adipokine primarily secreted by adipose tissue that enhances insulin sensitivity, reduces oxidative stress, and exerts anti-inflammatory effects. It plays a key role in regulating glucose levels and fatty acid oxidation. In the liver, it activates AMP-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor-α (PPAR-α) pathways, thereby suppressing gluconeogenesis, promoting lipid oxidation, and reducing hepatic steatosis [216].
Adiponectin levels are typically lower in individuals with obesity and metabolic disorders. This reduction is associated with the activation of IR and inflammatory processes, which may contribute to the development of MASLD [216]. In the context of liver diseases, serum adiponectin levels provide valuable insights into a patient’s metabolic status. Notably, adiponectin has been incorporated into biomarker models for the detection of MASH. In one study, the combination of cleaved CK-18, serum adiponectin, and resistin achieved a sensitivity of 95.45% and a specificity of 70.21%, suggesting that this panel may serve as a significant predictor for the diagnosis of MASH [217].
Moreover, several pharmacological studies have demonstrated that adiponectin receptor agonists (e.g., AdipoRon, ADP355) reduce steatosis, suppress inflammation, and improve insulin sensitivity in MASLD/MASH models [218]. These findings suggest that adiponectin may serve not only as a biomarker but also as a potential therapeutic target. However, some studies have reported elevated adiponectin levels in advanced liver disease and HCC, indicating that the effects of adiponectin may vary depending on the stage of liver pathology [219].
Hence, adiponectin is a key adipokine with a protective role in the pathophysiology of MASLD. Monitoring serum adiponectin levels may assist in evaluating disease progression, while therapies targeting adiponectin signaling pathways could represent promising strategies for the future management of MASLD.
Resistin: Resistin is an adipokine synthesized in adipocytes and other cell types and is associated with inflammation, IR, and metabolic diseases. It is considered to play a significant role in the development of MASLD and may be involved in the pathogenesis of hepatic steatosis and inflammation [213]. Resistin induces IR by inhibiting insulin receptor substrate 1 (IRS-1) phosphorylation via suppressor of cytokine signaling 3 (SOCS3) [214]. Particularly in conditions with increased IR, resistin may promote hepatic fat accumulation and inflammation [214]. Moreover, resistin has been suggested to aggravate hepatic inflammation and fibrosis by enhancing the production of pro-inflammatory cytokines [217].
Several studies have reported an association between elevated resistin levels and MASLD. Resistin may also play a crucial role in hepatic insulin desensitization, thereby accelerating the development of metabolic disorders. However, studies investigating the effects of resistin in MASLD have yielded conflicting results [220].
Further research is needed to better understand the relationship between resistin and MASLD. Nevertheless, current evidence suggests that resistin could represent a potential biomarker and therapeutic target in the progression of liver disease.
Visfatin: Visfatin, a 52-kDa adipokine that is highly conserved across all species, also known as nicotinamide phosphoribosyltransferase (NAMPT) and pre-B cell colony-enhancing factor 1 (PBEF-1), has several main sources, including adipocytes, lymphocytes, monocytes, neutrophils, hepatocytes, and pneumocytes. Among the various pathways affected by visfatin are oxidative stress response, apoptosis, lipid and glucose metabolism, IR, and inflammation, which likely play a role in the pathogenesis of MASLD. The expression of visfatin is regulated by several cytokines, including TNF-α, IL-6, and lipopolysaccharides, which are known to support IR. Moreover, increased visfatin levels have been found to be associated with atherosclerotic disease and coronary artery disease, which are among the leading causes of death in MASLD. Although several studies have evaluated the role of visfatin and its effects on hepatic steatosis, fibrosis, and inflammation in MASLD, the current evidence is inconclusive and inconsistent. According to the results of a meta-analysis including 14 studies, visfatin levels are not associated with MASLD, liver steatosis, liver fibrosis, lobar inflammation, or MASH [221].
Cell death markers
Caspase-cleaved keratin-18 (CK-18): Keratin 18 (K18) is one of the main intermediate filament proteins in hepatocytes, providing integrity to the cellular skeleton. Elevated hepatocyte apoptosis is well documented as a distinct feature of MASH [222]. During apoptosis, K18 proteins are cleaved by caspase enzymes at specific sites. These cleavages contribute to the reorganization of the cytoskeleton and the progression of apoptosis [223]. The resulting caspase-cleaved cytokeratin-18 fragments circulate in the peripheral blood of MASH patients, and their plasma/serum levels are associated with the severity of MASH [224]. Feldstein et al. reported that CK-18 fragments were significantly elevated in the blood of MASH patients and served as an independent predictor of MASH, detecting the presence of MASH with over 90% specificity and an 80% negative predictive value [78]. According to this study, the likelihood of having MASH increased by 30% for every 50 U/l increase in CK-18 plasma levels (OR = 1.3, 95% CI: (1.1, 1.4). This finding further supports the notion that circulating caspase-cleaved CK-18 fragments serve as a valuable diagnostic biomarker for MASH progression.
Additionally, in a study of patients who underwent bariatric surgery, CK-18 levels were found to indicate continued liver damage after surgery [223]. This suggests that CK-18 could be a potential biomarker for monitoring liver damage.
Furthermore, CK-18 levels have also been shown to be related to systemic inflammation markers. In a study of colorectal cancer patients, elevated CK-18 levels were associated with higher systemic inflammation marker levels [225]. This suggests that CK-18 might also be an important biomarker for assessing inflammation. These findings indicate that CK-18 is a crucial biomarker for evaluating oxidative stress and inflammation in MASLD. Current literature supports the use of CK-18 levels for monitoring disease progression and treatment response.
High mobility group box 1 (HMGB1): HMGB1 is a nuclear protein that plays essential roles within the cell, such as organizing genetic material and facilitating DNA repair. When released extracellularly, HMGB1 acts as an alarmin that modulates inflammation and immune responses. Therefore, the extracellular release of HMGB1 plays a critical role in the pathogenesis of various inflammatory diseases. In recent years, the association of HMGB1 with metabolic diseases has been increasingly investigated. HMGB1 protein levels are elevated particularly in liver diseases, especially in conditions such as MASLD [226].
In MASLD, the increased release of HMGB1 contributes to disease progression by enhancing inflammation. In this disease, characterized by obesity and hepatic steatosis, HMGB1 may trigger oxidative stress and inflammatory processes in hepatocytes. The release of HMGB1 can exacerbate cellular damage and inflammation, potentially leading to hepatitis and steatohepatitis [227].
HMGB1 is known to activate several signaling pathways that initiate and sustain inflammation. Among these are inflammatory cascades such as NF-κB and MAPK (mitogen-activated protein kinases). These signaling pathways lead HMGB1 to enhance the release of inflammatory cytokines, thereby increasing the level of inflammation in the liver. Additionally, it has been demonstrated that HMGB1 may contribute to the pathogenesis of MASLD by promoting insulin resistance [226].
The role of HMGB1 in MASLD is particularly associated with interactions between liver cells and immune cells. While HMGB1 contributes to the development of hepatic steatosis, it may also lead to the activation of Kupffer cells and the exacerbation of inflammation [227]. This condition emerges as a significant factor in disease progression. Moreover, HMGB1 levels rise in parallel with the severity of the disease, and thus, this protein presents a potential therapeutic target as one of the biomarkers of MASLD [228, 229].
In conclusion, HMGB1 stands out as a molecule that plays a significant role in the pathogenesis of MASLD by promoting inflammation. The increase in HMGB1 associated with liver damage may serve as an important biological marker to evaluate the severity and prognosis of the disease [227]. Therapeutic strategies targeting HMGB1 may offer a novel approach in the treatment of MASLD [228, 229].
Survivin: Survivin is an anti-apoptotic protein that exerts its effect by inhibiting caspase-3. In MASLD-related HCC, the expression of survivin is increased [224].
Soluble Fas ligand (sFASL): Soluble Fas ligand (sFASL) is the circulating form of membrane-bound Fas ligand, a member of the TNF superfamily, and plays a critical role in regulating apoptosis and immune responses. sFASL binds to the Fas receptor (CD95) on the surface of target cells, activating the caspase cascade and triggering programmed cell death. In the context of liver diseases, particularly conditions such as MASLD, dysregulation of apoptosis is considered a key mechanism contributing to hepatocyte damage, inflammation, and fibrogenesis [230].
In MASLD, hepatocyte apoptosis is a prominent feature that distinguishes simple steatosis from MASH. Elevated levels of sFASL have been observed in patients with MASLD, and this increase is thought to reflect enhanced apoptotic activity in hepatocytes. The rise in sFASL levels may serve both as an indicator of disease severity and as a mediator of liver injury by perpetuating inflammatory signaling and promoting further hepatocyte apoptosis [231].
Clinical studies have demonstrated that sFASL levels correlate with histological features of liver injury, particularly ballooning degeneration, lobular inflammation, and fibrosis. Moreover, sFASL has been shown to interact with immune cells such as T lymphocytes and macrophages, enhancing the production of pro-inflammatory cytokines and contributing to the chronic inflammatory milieu characteristic of MASLD [231, 232].
Due to its role in promoting hepatocyte apoptosis and inflammation, sFASL has emerged as a potential biomarker for monitoring disease progression in MASLD. Therapeutic strategies targeting the Fas/FasL signaling pathway may offer a novel approach to reduce hepatocyte damage and slow the progression of steatohepatitis and fibrosis.
New generation biomarkers and multi-omic approaches
MASLD has a globally increasing prevalence and has the potential to progress to its more severe form, MASH. During this transformation process, inflammation, hepatocyte damage, and fibrosis develop in the liver. While traditional biomarkers may be insufficient in the early stages of this process, new-generation biomarkers and multi-omic approaches hold the potential to contribute to the improvement of disease diagnosis and prognosis.
Multi-omic approaches (the combined evaluation of genomic, transcriptomic, proteomic, and metabolomic analyses) provide a better understanding of the underlying pathophysiology of MASLD. Recent studies have identified molecular biomarkers of liver damage through these approaches, revealing promising candidates for the early diagnosis of the disease [233, 234].
Fibroblast growth factor-21 (FGF21): Fibroblast growth factor-21 (FGF21) is a hepatokine produced by various tissues, primarily the liver, and plays a role in maintaining metabolic homeostasis. FGF21 is known for its effects on regulating glucose and lipid metabolism, increasing insulin sensitivity, and promoting energy expenditure. In the context of MASLD, FGF21 has been shown to be associated with hepatic steatosis and inflammation. It has been reported that FGF21 levels are significantly elevated in MASLD, particularly in MASH patients, and this increase may reflect hepatic stress and damage [235, 236].
Furthermore, FGF21 has been suggested as a potential biomarker for the diagnosis of MASLD and the assessment of disease severity. In histologically confirmed MASLD cases, FGF21 levels have been found to correlate with the degree of steatosis and ballooning degeneration [237, 238].
Therapeutically, FGF21 analogs are being explored as a new approach in the treatment of MASLD. Dual agonists combining FGF21 and GLP-1 receptor agonists have shown effectiveness in reducing hepatic steatosis and improving metabolic parameters. For example, a randomized controlled phase I study conducted on a new GLP-1/FGF21 dual agonist, HEC88473, demonstrated a significant reduction in liver steatosis after 5 weeks of treatment [239].
In light of this data, FGF21 stands out as an important hepatokine for both diagnostic and therapeutic purposes in MASLD, particularly for early diagnosis, monitoring disease progression, and targeting treatment [237].
Gut microbiota metabolites
In recent years, significant findings have been made regarding the role of gut microbiota metabolites in the pathogenesis of MASLD. Notably, trimethylamine N-oxide (TMAO) and bile acids are prominent metabolites in this context.
Trimethylamine N-oxide (TMAO): TMAO is a product formed when the gut microbiota metabolizes components such as choline and lecithin found in the diet. Although high levels of TMAO have been associated with CVDs, recent studies suggest that it is also linked to MASLD. TMAO is believed to contribute to the progression of the disease by increasing hepatic fat accumulation and triggering inflammation [240–242]. However, the mechanisms underlying this relationship remain unclear, and further studies are needed.
In the gut, Firmicutes bacteria convert dietary choline into trimethylamine (TMA); TMA is then oxidized in the liver by the enzyme flavin monooxygenase-3 (FMO3) to form trimethylamine N-oxide (TMAO). TMAO is a critical metabolite in microbiota dysbiosis that triggers hepatic inflammation. Foods rich in choline (e.g., eggs, red meat) increase TMAO production [241, 243–245]. Therefore, TMAO levels + microbiota analysis can be used to plan dietary interventions.
Bile acids
Bile acids are steroid acids synthesized from cholesterol in the liver and play a crucial role in fat digestion. Primary bile acids, such as cholic acid and deoxycholic acid, are directly synthesized from cholesterol in the liver, while secondary bile acids, such as lithocholic acid and ursodeoxycholic acid, are formed in the intestine as a result of bacterial metabolism [246, 247].
Bile acids regulate metabolic processes such as gluconeogenesis, lipogenesis, and inflammation by interacting with nuclear receptors like FXR (farnesoid X receptor) and TGR5 [248]. Specifically, FXR activation modulates glucose and lipid metabolism by reducing the expression of key enzymes involved in gluconeogenesis and lipogenesis. Dysregulation in the FXR and TGR5 signaling pathways leads to hepatocyte toxicity due to an increase in primary bile acids like cholic acid (CA) and chenodeoxycholic acid (CDCA), as well as a decrease in secondary bile acids such as deoxycholic acid (DCA) and lithocholic acid (LCA), which in turn triggers microbiota dysbiosis and MASLD pathogenesis [249]. Furthermore, bile acids are thought to play a role in modulating the progression of MASLD by affecting the gut microbiota [174, 180, 247, 250]. These findings suggest that regulation of bile acid metabolism is both a diagnostic and therapeutic target in MASLD.
Specific inflammatory biomarkers for early detection of MASLD
Chronic low-grade inflammation is a central feature in the pathogenesis of MASLD. While traditional inflammatory markers such as CRP and TNF-α reflect systemic inflammation, they lack specificity and sensitivity for early-stage MASLD. Recent research has identified more specific biomarkers that may serve as early indicators of hepatic inflammation and fibrogenesis in MASLD.
Among these, IL-6 has emerged as a key cytokine with both diagnostic and prognostic potential. Elevated IL-6 levels are associated with hepatic steatosis severity and correlate with insulin resistance and hepatic fat accumulation independent of BMI and other metabolic parameters [251]. Moreover, IL-1β has been shown to trigger hepatic Kupffer cells and hepatocyte injury in MASLD models, suggesting its possible role in early disease stages [252].
Another promising marker is soluble CD163 (sCD163), a macrophage activation marker that reflects hepatic macrophage activity. sCD163 levels are elevated in early MASLD and correlate with histological inflammation and fibrosis [253]. Similarly, CK-18 fragments, released during hepatocyte apoptosis, are elevated in MASLD patients and have been suggested as a non-invasive biomarker for MASH [254].
Emerging biomarkers such as MCP-1 and resistin are also under investigation. MCP-1 mediates immune cell recruitment and fibrogenesis, while resistin links obesity-associated inflammation with hepatic IR [255, 256].
Overall, these specific biomarkers provide a more nuanced view of the inflammatory milieu in MASLD and may enhance early detection, stratification, and monitoring of disease progression, especially when used in combination with imaging or omics-based tools.
Genetic and epigenetic biomarkers in MASLD
Heritability has a significant impact on several metabolic disorders, including obesity, IR, steatosis, and fibrosis – important factors contributing to the progression of MASLD [257, 258]. To fully understand the nature of complex and multifactorial diseases such as MASLD, it is important to consider the role of genetic and epigenetic factors as essential aspects of a broader context that includes clinical and environmental influences and their interactions when multiple metabolic pathways are involved [259].
SNPs as genetic biomarkers
Genetic biomarkers for MASLD are valuable tools for identifying individuals at increased risk of developing the disease for early detection and progression to more severe disease stages [257]. Several genes contribute to both susceptibility and progression of MASLD, with the most extensively studied genetic variants found in patatin-like phospholipase domain-containing 3 (PNPLA3), transmembrane 6 superfamily member 2 (TM6SF2), and membrane-bound O-acyltransferase domain-containing 7 (MBOAT7) [257].
PNPLA3 is mainly expressed in the liver and adipose tissue and encodes the enzyme adiponutrin. This enzyme is involved in the hydrolysis of TG into FFAs and glycerol and facilitates the mobilization of FFAs and energy utilization [260].
This enzyme has a dual activity: triacylglycerol lipase and acylglycerol O-acyltransferase. It is associated with lipid droplets in adipocytes and hepatocytes. It not only plays an important role in regulating TG levels but also hydrolyzes retinyl esters to retinol (vitamin A), thereby influencing vitamin A metabolism in hepatic stellate cells (HSC). Through its transacylase activity, it influences phospholipid metabolism [261].
One of the best-studied single nucleotide polymorphisms (SNPs) in the PNPLA3 gene is rs738409, in which cytosine (C) is replaced by guanine (G). This change in a nucleotide leads to an isoleucine-methionine substitution at position 148 (I148M) in the polypeptide chain [262]. The rs738409 G allele is the strongest genetic determinant of MASLD risk. It is associated with a higher risk of progression of steatosis to MASH, fibrosis, and cirrhosis. It carries an increased risk of developing HCC [263]. This polymorphism impairs the lipase activity of adiponutrin and impairs TG hydrolysis, leading to an accumulation of TG in hepatocytes and consequently to steatosis. This also favors the deposition of unsaturated fatty acids and toxic lipid intermediates. Lipid overload triggers inflammation and oxidative stress, leading to mitochondrial and endoplasmic reticulum stress [262, 264]. Homozygotes for the G allele tend to have a higher risk of more aggressive progression from MASLD to cirrhosis and an increased risk of HCC [263, 264].
Several other SNPs in the PNPLA3 gene that may be associated with MASLD have also been investigated.
The rs2294918 G>A polymorphism in the PNPLA3 gene leads to a substitution of glutamic acid (E) with lysine (K) at position 434 of the adiponutrin protein (E434K). Carriers of the A allele rs2294918 have reduced hepatic PNPLA3 messenger ribonucleic acid (mRNA) and protein levels, suggesting that this variant reduces PNPLA3 expression without affecting adiponutrin enzymatic activity [265]. A potential interaction between the rs2294918 G>A and rs738409 C>G variants has been proposed to influence the risk of progression to MASLD. The presence of the rs2294918 A allele decreased adiponutrin expression, thereby reducing the effects of the rs738409 G allele on predisposition to steatosis and liver damage [265].
The polymorphism rs6006460 G>T in the PNPLA3 gene leads to a substitution of serine (S) with isoleucine (I) at position 453 of the adiponutrin protein (S453I). The presence of the rs6006460 T allele appears to have a protective effect against the accumulation of TG in hepatocytes, which could reduce the risk of developing MASLD [262]. This effect is independent of other known risk factors, suggesting a direct influence of the S453I variant on hepatic lipid metabolism.
PNPLA3 polymorphisms are potential biomarkers for screening and risk stratification for the development and progression of MASLD. In addition to the combined effects of PNPLA3 variants rs738409 I148M and rs2294918 E434K on the pathogenesis of MASLD, they are also considered to be independent genetic determinants of MASLD [265].
The transmembrane 6 superfamily member 2 (TM6SF2) protein is mainly expressed in the liver and plays a key role in the maintenance of lipid homeostasis by controlling the secretion of lipids from the liver [266]. The activity of TM6SF2 is required for the normal secretion of TG-rich lipoproteins such as very low-density lipoproteins (VLDL). Decreased expression of TM6SF2 can lead to decreased VLDL secretion and increased TG content in the liver as well as lipid droplet formation [266, 267]. TM6SF2 may function as an enzyme in cholesterol metabolism [268]. Overexpression of TM6SF2 led to an increase in total cholesterol and LDL cholesterol (LDL-c), and a decrease in HDL-c, while downregulation of TM6SF2 may lead to a decrease in total cholesterol.
A notable single nucleotide polymorphism (SNP) in the TM6SF2 gene is rs58542926, in which guanine (G) is replaced by adenine (A) at position 499 of the coding region (c. 499 G>A). This change leads to a substitution of glutamic acid (E) with lysine (K) at position 167 (E167K, p.Glu167Lys) [267]. Homozygous carriers of the A allele have a higher content of polyunsaturated TG in the liver and are more likely to develop MASH than G allele homozygotes [269].
Carriers of the A allele of TM6SF2 not only exhibit hepatic steatosis and a decrease in circulating TG, LDL-c, and total cholesterol levels but can also show endoplasmic reticulum stress, which can lead to inflammation and fibrosis and thus to disease progression [270].
In addition to lipid accumulation and inflammation, this variant influences the development and progression of MASLD by stimulating oxidative stress. In the study by Sun and colleagues, the A allele of this variant was found to be associated with increased levels of MDA and ROS, which can impair cell membrane fluidity and contribute to liver damage [271]. The E167K variant enhances the interaction between TM6SF2 and the PNPLA3 variant I148M, which inhibits the function of PNPLA3 in mediating the transfer of polyunsaturated fatty acids (PUFAs) from TG to phosphatidylcholine, leading to increased accumulation of polyunsaturated TG in the liver and exacerbation of liver injury [271].
Membrane bound O-acyltransferase domain-containing 7 (MBOAT7) encodes a membrane-bound enzyme that is involved in the biosynthesis and restructuring of phosphatidylcholine (PC) and other phospholipids and in particular catalyzes the O-acylation of lysophospholipids. This enzymatic activity is crucial for maintaining the fluidity and integrity of the cell membrane [272].
Genetic variants in MBOAT7 have been identified as a significant risk that may contribute to the abnormal formation of lipid droplets that promote liver injury, inflammation, and the development and progression of MASLD [273].
The best-studied and characterized genetic variant in MBOAT7 is the polymorphism rs641738, which leads to a substitution of cytosine (C) with thymine (T) at position 641738 in the gene. Rs641738C>T is associated with reduced MBOAT7 mRNA [274] and protein expression [273]. The T allele reduces MBOAT7 enzyme activity, impairs phospholipid remodeling, and promotes hepatic lipid accumulation. Carriers of the T allele have increased production of diacylglycerols and ceramides (lipotoxic intermediates), which accumulate in liver cells and trigger inflammation [272, 274]. The inability to efficiently remodel phospholipids also disrupts the balance between cholesterol and phospholipids in lipoproteins such as VLDL. These carriers have been found to have higher ALT activity, indicating damage to hepatocytes. They have a higher risk of developing severe forms of liver disease, including advanced fibrosis and cirrhosis [275]. Heterozygous (C/T) individuals have a moderate risk, while homozygous (T/T) individuals face severe liver complications.
In addition to the well-characterized rs641738 variant, the rs8736 variant in the MBOAT7 gene was also identified and investigated in connection with MASLD. This SNP leads to a substitution of cytosine (C) with thymine (T) at the position 54173495. Similar to individuals with the rs641738 TT genotype, homozygous individuals with the T-risk allele (rs8736 TT) have lower MBOAT7 expression [273, 276]. They respond less strongly to TLR4 stimulation than individuals with the CC genotype. In addition, reduced MBOAT7 expression in TT carriers led to increased levels of inflammatory cytokines (TNF-α and MCP-1) after stimulation with TLR2, TLR4, and TLR9 compared to controls [276].
Integrative genome-wide association studies (GWAS) and meta-analyses have identified the genetic variants in PNPLA3, TM6SF2 and MBOAT7 as key factors associated with increased liver injury risk and severity of MASLD [277, 278]. Taken together, integrative analyses show that these three loci modulate MASLD risk via different biological pathways: PNPLA3 via lipid droplet remodeling, TM6SF2 via impairment of hepatic lipid export, and MBOAT7 via membrane phospholipid turnover [278]. In the multivariable model PNPLA3 p.I148M and TM6SF2 p.E167K were associated with steatosis, while PNPLA3 p.I148M and MBOAT7 Rs641738C>T were significantly associated with the development of liver fibrosis. Their additive effects are evident in polygenic risk models, with PNPLA3 being the major contributor [279]. A recent meta-GWAS study showed that individuals in the top decile of genetic risk (across all loci of PNPLA3, TM6SF2, and MBOAT7) have a significantly elevated risk of MASLD, cirrhosis, and HCC (OR up to 6-fold) [280].
Polygenic risk scores (PRS) incorporating PNPLA3, TM6SF2, and MBOAT7 can predict the likelihood of developing liver fat accumulation, inflammation and fibrosis and help to identify individuals at high risk of liver disease progression, particularly in situations where liver biopsy is not possible or practical [281]. The clinical utility of these SNPs in genetic models could improve diagnosis and risk stratification in patients with obesity and T2D with MASLD.
miRNAs as epigenetic biomarkers
In addition to important epigenetic markers such as DNA methylation, histone acetylation and long non-coding RNAs, micro-ribonucleic acids (miRNAs) are the most studied.
Over the last few decades, research in the field of miRNAs has expanded considerably, positioning them as potentially promising epigenetic biomarkers. As small, non-coding RNA molecules, typically 19 to 25 nucleotides in length, they play a crucial role in post-transcriptional gene regulation [282] and influence various physiological and pathological processes such as cell growth, differentiation, developmental processes, aging, metabolism, apoptosis, and oncogenesis [283].
They bind to complementary target messenger RNA (mRNA) predominantly in the 3′-untranslated region, leading either to its degradation or to inhibition of protein synthesis, depending on the degree of sequence complementarity between the miRNA and the mRNA [282]. If there is a perfect match, the mRNA is cleaved and subsequently degraded. Conversely, an incomplete base pairing leads to suppression of translation. Each miRNA is capable of affecting multiple mRNA transcripts, so it can potentially regulate hundreds of different genes [284]. Although miRNAs are primarily known for repressing translation, some of them can also activate gene expression under certain conditions. This activation can occur through direct interaction with the target mRNA to increase its stability or promote translation [284].
In addition to their role in gene regulation in cells, miRNAs are associated with various biological molecules, including proteins and lipoproteins, which increases their stability in the bloodstream [282]. In general, miRNAs are either bound to Argonaute-2 proteins (AGO2) or encapsulated in extracellular vesicles such as exosomes, microvesicles, or apoptotic bodies [285], which protects them from enzymatic degradation and ensures their stability. This makes them ideal candidates for non-invasive diagnostic tools. In particular, dysregulation of miRNA expression has been observed in liver diseases ranging from steatosis to HCC, making it particularly useful for the assessment of disease progression and response to treatment [282, 286].
As already mentioned, the early stages of MASLD are characterized by the interplay of dyslipidemia, inflammation, and oxidative stress. In this section, the most important miRNAs associated with these pathological mechanisms are presented.
MiRNA-122 (miR-122) is one of the best-characterized liver-specific miRNAs, and it accounts for about 70% of all miRNAs in hepatocytes [287]. It plays a crucial role in liver homeostasis and lipid metabolism. In addition, this miRNA has been shown to be a regulator of inflammation and tumorigenesis [288].
miR-122 is important for the maintenance of liver tissue identity and the regulation of hepatocyte differentiation. It stimulates hepatocyte proliferation and maturation by modulating the expression of hepatocyte nuclear factor 6 (HNF6) [289], influences cell growth and survival pathways in liver cells by modulating insulin-like growth factor 1 receptor (IGF1R), and modulates cell survival and growth through phosphatase and tensin homolog (PTEN) [290].
As an important regulator of lipid metabolism, miR-122 influences the synthesis, oxidation, and storage of cholesterol and fatty acids [291, 292]. The role of miR-122 in the early stages of MASLD is still controversial. It has been shown that miR-122 has a dual influence on hepatic lipid metabolic pathways, a pro- and an anti-steatotic direction [290–292].
miR-122 can promote hepatic lipogenesis by inhibiting the liver kinase B1 (LKB1)/5′ adenosine monophosphate-activated protein kinase (AMPK) signaling pathway through direct targeting of sirtuin 1 (SIRT1) [291]. This inhibition leads to reduced activation of AMPK, resulting in increased lipid synthesis and accumulation in the liver. Conversely, miR-122 has been shown to inhibit lipid droplet formation and hepatic TG accumulation by enhancing the farnesoid X receptor and small heterodimer partner (FXR-SHP) signaling pathway, resulting in decreased lipid synthesis and storage in hepatocytes [292]. Furthermore, miR-122 decreases the expression of lipogenic enzymes, fatty acid synthase (FAS), and acetyl-CoA carboxylase (ACC), thereby decreasing TG synthesis and VLDL secretion and increasing β-oxidation of fatty acids [290, 293]. These results suggest that miR-122 exerts a protective effect against the development of MASLD.
miR-122 also plays a dual opposing role in inflammation. In MASLD, elevated miR-122 levels increase inflammation by activating the nuclear factor kappa B (NF-κB) signaling pathway, leading to the production of TNF-α, IL-6, and other proinflammatory cytokines that contribute to inflammation in the liver [294]. It also regulates hepatic inflammatory responses by directly targeting a member of the NF-κB family, RELB. In the livers of miR-122 knockout (KO) mice, levels of proinflammatory chemokines and RelB were elevated. The observed increase in RelB likely contributes to the increased expression of chemokines (Ccl2, Ccl4, Ccl20, Cxcl2, and Cxcl10) in the liver [295].
Elevated levels of miR-122 are associated with increased oxidative stress in liver cells. miR-122 directly inhibits forkhead box protein O3 (FOXO3) expression and impairs activation of the antioxidant enzyme SOD in cultured hepatocytes of the HFD-induced MASLD mouse model, leading to increased oxidative stress in liver cells [296]. This increased oxidative stress contributes to hepatocyte apoptosis and triggers inflammatory responses that promote fibrogenesis in MASLD.
In MASLD, the expression of miR-122 changes during the course of the disease. It is initially upregulated in steatosis but decreases during the transition to MASH and fibrosis. However, its serum levels correlate positively with the severity of steatosis, making it a promising diagnostic tool. In contrast, knockout models show that its deletion leads to steatosis, fibrosis, and the development of HCC, while its restoration reverses these effects [290].
miR-34a is less highly expressed in the liver than miR-122 but is upregulated in the liver of MASLD patients compared to controls [296, 297]. The expression of miR-34a in the liver is closely related to the severity of MASLD, and its levels appear to increase with disease progression [290, 296, 297]. It influences the progression of MASLD by increasing lipid uptake and lipid synthesis while promoting inflammation and cell death.
It is known to suppress the activity of several important genes involved in the progression of MASLD, including hepatocyte nuclear factor 4α (HNF4α), peroxisome proliferator-activated receptor α (PPARα), and SIRT1 [297].
In patients with MASH and in mice fed a high-fat diet (HFD), miRNA-34a was found to play a role in the regulation of hepatic VLDL metabolism through its interaction with HNF4α [298]. Decreased expression of HNF4α by miR-34a in turn decreases cAMP-responsive element binding protein H (CREBH), which impairs VLDL assembly and secretion, leading to lipid accumulation in hepatocytes and possibly contributing to hepatic steatosis [299].
MiR-34a represses the expression of PPARα, leading to reduced transcription of its target genes involved in fatty acid β-oxidation. This downregulation affects genes such as carnitine palmitoyltransferase 1 (CPT1), carnitine palmitoyltransferase 2 (CPT2), and fatty acid transport proteins [298]. Consequently, impaired PPARα signaling disrupts mitochondrial fatty acid β-oxidation, leading to lipid accumulation in hepatocytes and possibly accelerating the progression of MASLD [297]. Experimental studies have shown that inhibition of miR-34a in mice fed an HFD increases PPARα expression and AMPK activation, which promotes fatty acid β-oxidation and reduces TG accumulation in the liver [300].
miR-34a inhibits SIRT1 directly by binding to its 3′-untranslated region and indirectly by reducing NAD+ availability through suppression of the enzyme nicotinamide phosphoribosyltransferase (NAMPT) [301]. By inhibiting the same transcription factor, miR-34a has been associated with an increase in cholesterol production in MASLD patients by maintaining 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase (HMGCR) in its active form and decreasing the activation of AMPK, a key enzyme in energy consumption [298].
In addition, lower SIRT1 levels trigger the activation of apoptosis-related genes such as P66SHC and P53, which increases the susceptibility of hepatocytes to oxidative stress and programmed cell death. In experimental models, blocking miR-34a restores SIRT1 activity, leading to increased TG degradation, increased β-oxidation of fatty acids, and consequently a reduction in fat accumulation in the liver [302]. By directly downregulating SIRT1, miR-34a leads to increased synthesis of inflammatory cytokines, including IL-6, TNF-α, IL-1β, and IL 8, and oxidative stress proteins, including MCP-1, vascular cell adhesion protein 1 (VCAM1), and MDA, and decreased levels of the anti-inflammatory cytokine IL-10 [297, 303].
MiR-21 is widely expressed in human tissues and is upregulated mainly in non-parenchymal liver cells such as HSCs, inflammatory cells, and biliary cells [290, 304]. It is known to be dysregulated in diseases such as cancer, inflammation, fibrosis, and MASLD [305].
Studies using germline and hepatocyte-specific conditional miR-21 knockout mice have shown that miR-21 remains inactive in hepatocytes under normal physiological conditions [306]. However, upon HFD treatment, miR-21 was activated and promoted lipid accumulation and IR in hepatocytes through its influence on key metabolic enzymes involved in glucose and lipid metabolism [306]. Studies suggest that miR-21 affects hepatic lipid metabolism by suppressing its direct target, PPARα, in human liver cells [305]. By downregulating PPARα, miR-21 in hepatocytes together with miR-34a could promote TG accumulation in the early stages of MASLD but also induce inflammation and fibrosis progression [290, 307]. Inhibition of miR-21 attenuates steatosis through activation of PPARα. In addition, miR-21 inhibits fatty acid binding protein 7 (FABP7) [308], which specifically promotes fatty acid uptake and lipid accumulation.
In contrast, miR-21 regulates TG and cholesterol metabolism in the MASLD in vitro model via a different mechanism. This effect is achieved by inhibiting HMGCR expression, its direct target. Enhanced expression of miR-21 resulted in a significant decrease in HMGCR mRNA and protein expression in HepG2 cells, leading to a reduction in TG and cholesterol levels [309].
The results of the study by Lu et al. identified the signal transducer and activator of transcription, STAT3, an inflammation-responsive transcription factor, as a target of miR-21-5p. They showed that inhibition of miR-21-5p can downregulate the expression of the IL-6 receptor (IL6R)/STAT3 signaling pathway in ulcerative colitis and apoptosis-associated proteins to alleviate inflammation and reduce apoptosis in RAW264.7 cells [310]. The upregulation of miR-21 in macrophages, the key cells of the inflammatory response in MASLD, has been associated with a systemic inflammatory response and increased production of proinflammatory cytokines such as IL-1β, TNFα, and IL-6 [311].
In contrast to these findings, other studies suggest that miR-21 is important for the progression rather than the onset of MASLD [305]. While its hepatic expression remains relatively unchanged in the early stages of MASLD [290, 305], miR-21 levels were significantly elevated in patients with MASLD compared to healthy controls and individuals with steatosis [312]. In some studies, no significant difference in serum levels of miR-21 was found between MASLD patients with fibrosis and healthy individuals, and its levels did not correlate with the severity of steatosis, fibrosis stage, and inflammation, in contrast to miR-34a and miR-122 [290].
miR-192 is highly expressed in the liver, where it regulates liver development, cellular transdifferentiation, and energy metabolism [313].
miR-192 was mainly observed in HSC and played a crucial role in inhibiting their activation, proliferation, and migration. Increased expression of miR-192 was found in MASLD patients compared to healthy controls, and it correlates with the severity of MASLD [271]. Like miR-122, the serum concentration of miR-192 is also increased in patients with MASH [290, 314].
The study by Lin et al. confirmed that miR-192 directly acts on the 3′UTR of sterol regulatory element binding transcription factor 1 (SREBF1) and contributes to the dysregulation of lipid homeostasis in hepatocytes. When miR-192 is absent or inhibited, SREBF1 expression is increased, leading to enhanced de novo lipogenesis and promoting the development of MASLD [315].
miR-192 was identified as a negative regulator of lipid synthesis in hepatocytes [316]. Inhibition of miR-192 leads to increased expression of stearoyl-CoA desaturase 1 (SCD-1), the enzyme that catalyzes the biosynthesis of monounsaturated fatty acids [317], resulting in increased TG accumulation. Conversely, overexpression of miR-192-5p suppresses SCD-1 levels, thereby reducing lipid accumulation in hepatocytes. The same role of miR-192-5p has also been demonstrated in other signaling pathway. It has been shown to target Yin Yang 1 (Yy1), a transcription factor that influences the expression of FAS [318]. By inhibiting Yy1, miR-192-5p decreases FAS levels, leading to lower TG synthesis in the liver. This suggests that miR-192-5p serves as a protective regulator against hepatic steatosis in MASLD by modulating the SCD-1 and Yy1/FAS signaling pathways.
Elevated levels of circulating miR-192 have been associated with increased hepatic inflammatory activity in MASLD. In MASLD, hepatocytes undergoing lipotoxic injury release exosomes enriched in miR-192. These exosomes are internalized by macrophages, leading to their activation into a pro-inflammatory M1 phenotype [319]. Remarkably, serum levels of miR-192 correlated positively with hepatic inflammatory activity in patients with MASLD, highlighting its role in disease pathology.
MiR-155, which is expressed in hepatocytes, Kupffer cells and immune cells, plays a role in various biological processes [297]. MiR-155 is associated with the progression of MASLD, is upregulated in mouse models of MASH-like liver injury, and correlates with disease severity [320].
It is downregulated in the liver and blood of MASLD patients compared to healthy individuals. Liver X receptor α (LXRα) is a direct target of miR-155 [321]. Decreased miR-155 can activate the LXRα/sterol regulatory element binding protein 1c (SREBP-1c) and LXRα/FAS signaling pathways, contributing to cholesterol and TG synthesis in the liver [297]. Moreover, miR-155 may influence fatty acid accumulation by targeting CCAAT/enhancer-binding protein β (C/EBPβ) in HepG2 cells. Overexpression of miR-155 decreases C/EBPβ levels, thereby reducing the accumulation of fatty acids within lipid droplets in the liver [322]. The pro-steatotic and anti-steatotic effects of miR-155 may be achieved through the same signaling pathways by modulating protein expression of SREBP-1c and FAS. These contrasting results may be due to different stages of MASLD in the mouse models, suggesting that miR-155 may act as a protective feedback mechanism in the early stages of MASLD [297].
miR-155 could have an important function in the regulation of inflammation. The research findings of Csak et al. suggest that miR-155 deficiency exacerbates inflammatory responses and leads to increased levels of cytokines and chemokines such as TNF-α and MCP-1 in methionine-choline-deficient diet-induced steatohepatitis. Furthermore, miR-155 deficiency led to a reduction in steatosis and fibrosis in MASH despite the presence of significant inflammation. These observations suggest that miR-155 affects fibrosis through mechanisms that are at least partially independent of its effects on inflammation in the liver [320].
MiR-375 exerts a multifunctional role as a regulator of the expression of several genes related to lipid metabolism. The role of miR-375 includes modulation of lipid metabolism, inflammation, and liver fibrosis, making it a potential therapeutic target and biomarker for MASLD. It is mainly expressed by the pancreas, but the brain and spinal cord also synthesize it [323].
Decreased expression of miR-375 upregulated the expression of adiponectin, which in turn inhibited lipid accumulation and stimulated β-oxidation of fatty acids. In addition, the downregulation of miR-375 leads to increased expression of adiponectin receptor 2 (AdipoR2), which enhances adiponectin signaling and has similar effects. In palmitic acid (PA)-induced human HCC cells, HepG2, leptin levels, and concentrations of inflammatory cytokines, including TNF-α and IL-6, were decreased [324].
Indirectly, miR-375 may influence the development of MASLD by interfering with lipid metabolism in adipocytes. In 3T3-L1 adipocytes, miR-375 upregulates the expression of genes related to lipid metabolism, including glycerol-3-phosphate acyltransferase, mitochondrial (GPAM), C/EBPα, PPARγ, and others, contributing to TG synthesis and accumulation in lipid droplets [325].
In human aortic endothelial cells, decreased expression of miR-375 significantly reduced the expression of IL-6, IL-1β, TNF-α, p-IκBa/IκBα, ROS, and NOX-4, and attenuated oxidative stress and inflammation [323, 326].
The role of miR-126 in the pathogenesis of MASLD is complex. Its serum expression levels did not differ between MASLD patients and control subjects, but decreased circulating miR-126 was associated with severe disease in men with MASLD [327]. In another study, significantly increased miR-126 levels were found in the MASLD group compared to controls. The levels were significantly associated with steatosis, liver stiffness, and liver fat content. In particular, the combination of miR-126 levels with leptin concentrations showed excellent diagnostic accuracy in detecting steatosis [328].
miR-126 negatively regulates lipid metabolism by regulating genes involved in lipid synthesis and uptake. Inhibition of miR-126 resulted in increased lipid synthesis accompanied by increased levels of FAS, acyl-CoA synthetase long-chain family member 1 (ACSL1), and insulin-induced gene 1 (Insig1). Conversely, overexpression of miR-126 decreased lipid content and reduced the levels of FAS and Insig1 [329].
MiR-126-3p has been shown to increase lipid accumulation in 3T3-L1 cells via different mechanisms by inducing the expression of CCAAT/enhancer-binding protein α (C/EBPα) [330].
No data have been published on the role of miR-126 in oxidative stress and inflammation in the liver of MASLD patients.
MicroRNA profiling in MASLD detection
An innovative approach to exploit the potential of miRNAs as biomarkers for MASLD is the profiling of miRNAs from serum extracellular vesicles (EVs). These EVs protect miRNAs from degradation and may reflect pathological changes in tissues such as the liver [331]. EV-derived miRNAs have the advantage that they are stable in the blood and resistant to ribonuclease degradation, which increases their suitability for clinical application. A recent study by Hu et al. investigated miRNA profiles in serum EVs from MASLD patients. Using an efficient method employing wheat germ agglutinin (WGA)-coupled magnetic beads for EV isolation, they identified three significantly elevated miRNAs – miR-574-3p, miR-542-3p, and miR-200a-3p – in MASLD patients compared to healthy controls. These miRNAs showed promising diagnostic accuracy, especially miR-574-3p and miR-200a-3p, based on ROC curve analysis [332].
Tobaruela-Resola et al. [333] conducted a longitudinal study in which they investigated how circulating miRNAs respond to dietary interventions in overweight or obese individuals with MASLD. The researchers monitored miRNA expression over 6, 12, and 24 months while patients followed a weight-loss diet. Specific miRNA panels correlated with the resolution of MASLD. After 12 months, a panel of miR-29b-3p, miR-122-5p, and miR-151a-3p in combination with BMI showed excellent diagnostic power (AUC = 0.85) for MASLD. Even after 24 months, miR-21-5p, miR-151a-3p, and BMI effectively discriminated between patients in whom steatosis had regressed and those in whom it had not. Interestingly, miR-122-5p, a liver-enriched miRNA commonly associated with liver injury, was consistently associated with liver enzymes (ALT, AST, and GGT) and steatosis severity, highlighting its utility as a dynamic biomarker responsive to intervention [333].
Zhang et al. [334] extended the utility of miRNA profiling to pediatric MASLD and identified 36 miRNAs with altered expression levels in affected children. In particular, miR-122-5p, which is already established as a critical hepatic biomarker, was upregulated in MASLD serum, consistent with studies in adults. The other significantly elevated miRNAs included miR-192-3p, miR-27a, miR-335-5p, and miR-34a, all of which have previously been linked to liver fat accumulation, insulin resistance, and inflammation. Importantly, this pediatric-focused study showed that miRNA alterations start early and are not restricted to adult populations, suggesting that miRNA panels could help with early detection and prevention strategies in adolescents.
DNA methylation in MASLD detection
DNA methylation has been shown to be an important epigenetic mechanism linking genetic and environmental factors to the development and progression of MASLD [335]. Differential methylation, particularly in relation to active demethylation pathways, and expression patterns of genes regulating lipid metabolism, inflammation, and fibrosis appear to be closely linked to the development and exacerbation of MASLD, from steatosis to more severe forms such as MASH and fibrosis [335, 336].
During methylation, methyl groups are usually attached to the DNA, especially to the cytosines that precede the guanines (CpG sites), resulting in 5-methylcytosine (5mC). This change often leads to genes being silenced as their chromatin structure is less accessible. On the other hand, DNA demethylation can occur passively during DNA replication or actively through enzymatic processes. In active demethylation, 5mC is converted into oxidized intermediates such as 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxycytosine (5caC), which can alter gene expression and cell function [337]. Different methylation patterns correlate with MASLD progression, but the exact regulatory mechanisms are not yet fully understood. Some evidence, including animal and human studies, suggests that active demethylation intermediates such as 5hmC and 5fC may be a better indicator of liver fibrosis risk than total 5mC levels [335]. Individuals with MASLD and moderate to high fibrosis risk had progressively higher global 5hmC and 5fC levels. The presence of fibrosis was negatively associated with SOCS3 gene promoter methylation and positively associated with mRNA expression of SOCS3, TXNIP, IL-6, and MCP-1, which are involved in oxidative stress, inflammation, and fibrogenesis [335].
Hypomethylation of the PPARγ gene, involved in lipid metabolism, increases its expression and stimulates steatosis development [338]. Loss of PPARα function in hepatocytes initiates DNA hypermethylation of genes related to fatty acid, bile acid, and steroid hormone receptors, including PPARα, leading to progression of disease [339].
In a targeted DNA methylation study focusing on five genes involved in fibrogenesis modulation, the results showed hypomethylation of pro-fibrogenic genes (TGFβ1 and PDGFα) and hypermethylation of anti-fibrogenic genes (PPARα and PPARδ) in advanced MASLD compared to mild cases, suggesting that the methylation status of specific CpGs could potentially predict the severity and progression of MASLD to liver fibrosis [340].
Strengths and limitations
This review offers a comprehensive and up-to-date synthesis of current knowledge regarding biomarkers relevant to the early detection and progression of MASLD. One of the key strengths of this study is its integrative approach, combining oxidative stress, inflammatory, genetic, and epigenetic biomarkers with insights from multi-omics technologies and artificial intelligence applications. Additionally, the inclusion of recently approved therapies and ongoing clinical trials enhances the translational relevance of the review.
However, several limitations should be acknowledged. First, despite efforts to include the most recent literature, the field of MASLD is rapidly evolving, and emerging biomarkers or therapeutics may not yet be fully represented. Second, as a narrative review, this study is inherently limited by potential selection bias and lacks the methodological rigor of a systematic review. Lastly, some biomarkers discussed in this review are still under investigation and lack validation in large, diverse cohorts, which limits their immediate clinical applicability.
Conclusion and future perspectives
MASLD has a globally increasing prevalence with the potential to progress to its more severe form, MASH, and is an independent risk factor for cardiometabolic disorders. Oxidative stress and inflammation, as interdependent processes, play pivotal roles in the pathogenesis of MASLD. Moreover, heritability has a significant impact on MASLD. Hence, to fully understand the nature of complex and multifactorial diseases such as MASLD, it is important to consider the role of genetic and epigenetic factors as essential aspects of a broader context that includes clinical and environmental influences and their interactions when multiple metabolic pathways are involved. Finding a key that will not only alleviate the oxidative-inflammatory burden on the liver and reverse steatosis but also open the door to reducing cardiometabolic morbidity and mortality is of utmost importance. Traditional biomarkers are usually insufficient in the early stages of this process. By detailing the clinical applications of lipid peroxidation products, protein/DNA damage markers, reactive oxygen species, and genetic and epigenetic biomarkers, we aimed to contribute to the development of non-invasive diagnostic approaches in MASLD. Blood mRNA panels are emerging as promising non-invasive biomarkers that detect changes in gene expression indicative of liver inflammation, oxidative stress, dyslipidemia, fibrosis, and cirrhosis. These transcriptomic signatures can help to identify disease stages and progression and represent a potential alternative to liver biopsy for diagnosis and monitoring.
Among the biomarkers discussed, some are approaching clinical utility and have shown promise for bedside application. For example, serum 8-OHdG, ox-LDL, and CYP2E1 are being evaluated as non-invasive indicators of oxidative stress in MASLD. Notably, serum-based fibrosis scores (e.g., ELF test incorporating HA, PIIINP, TIMP-1) have already gained FDA breakthrough designation and are integrated into ongoing phase II/III clinical trials for MASLD drug development [341]. Inflammatory markers such as TNF-α and IL-6 are also under active investigation, particularly in studies evaluating anti-cytokine therapies (e.g., infliximab, tocilizumab) [342]. Additionally, transcriptomic biomarkers such as miR-122 are being explored in early-phase trials for risk stratification and treatment monitoring [343]. To facilitate the clinical integration of emerging biomarkers in MASLD diagnosis and management, a flowchart summarizing the proposed stepwise implementation – ranging from initial clinical assessment to biomarker-guided monitoring – is presented in Figure 4. However, before these biomarkers can be adopted into routine clinical use, further standardization, cost-effectiveness evaluation, and regulatory validation are required.
In the future, combined diagnostic markers, targeted therapies, and multi-omic-based patient stratification that simultaneously target these two pathogenic axes – oxidative stress and inflammation – could have the potential to radically change clinical outcomes. Genetic and epigenetic biomarkers are certainly an inevitable part of the innovative technologies in MASLD management, i.e., diagnosis, risk assessment, and stratification of the disease. Regardless of all the above, all patients diagnosed with MASLD should be strongly encouraged to adopt primarily non-pharmacological interventions, with particular emphasis on promoting a healthy lifestyle, maintaining a balanced diet, and engaging in regular physical activity in order to reduce the risk of MASLD progression and associated adverse outcomes.