Introduction
Ulcerative colitis (UC) has become a global disease with approximately 5 million patients worldwide [1]. UC is a complex chronic inflammatory disorder of the colon with an immune-mediated etiology. Several risk factors have been implicated, including dietary habits, pharmaceutical exposures, and lifestyle elements that can modulate the host microbiome or antigenic immune responses [1, 2].
UC features complex pathologies including epithelial barrier disruption, dysregulated immune responses, and microbiome alterations [1]. Histologically, active UC shows neutrophil infiltration, crypt abscesses, goblet cell depletion, and mucosal ulceration, while the quiescent phase exhibits glandular distortion, submucosal fibrosis, and lymphatic dilation [3]. Among these multifaceted pathologies, aberrant angiogenesis – the formation of new blood vessels – has emerged as a critical driver of inflammation and tissue damage [4]. Dysfunctional vasculature perpetuates leukocyte infiltration, disrupts mucosal oxygenation, and contributes to the vicious cycle of inflammation [4, 5]. At present, conventional therapies for UC include biologic therapy, herbal therapy, and fecal microbiota transplantation [6]. Furthermore, diet and exercise, which promote recovery from hyperlipidemia, diabetes, and hypertension in older patients, are considered essential adjunctive therapies for UC [7, 8]. Among them, natural compounds from herbs are attracting increasing attention for development of new therapeutic agents for UC [2].
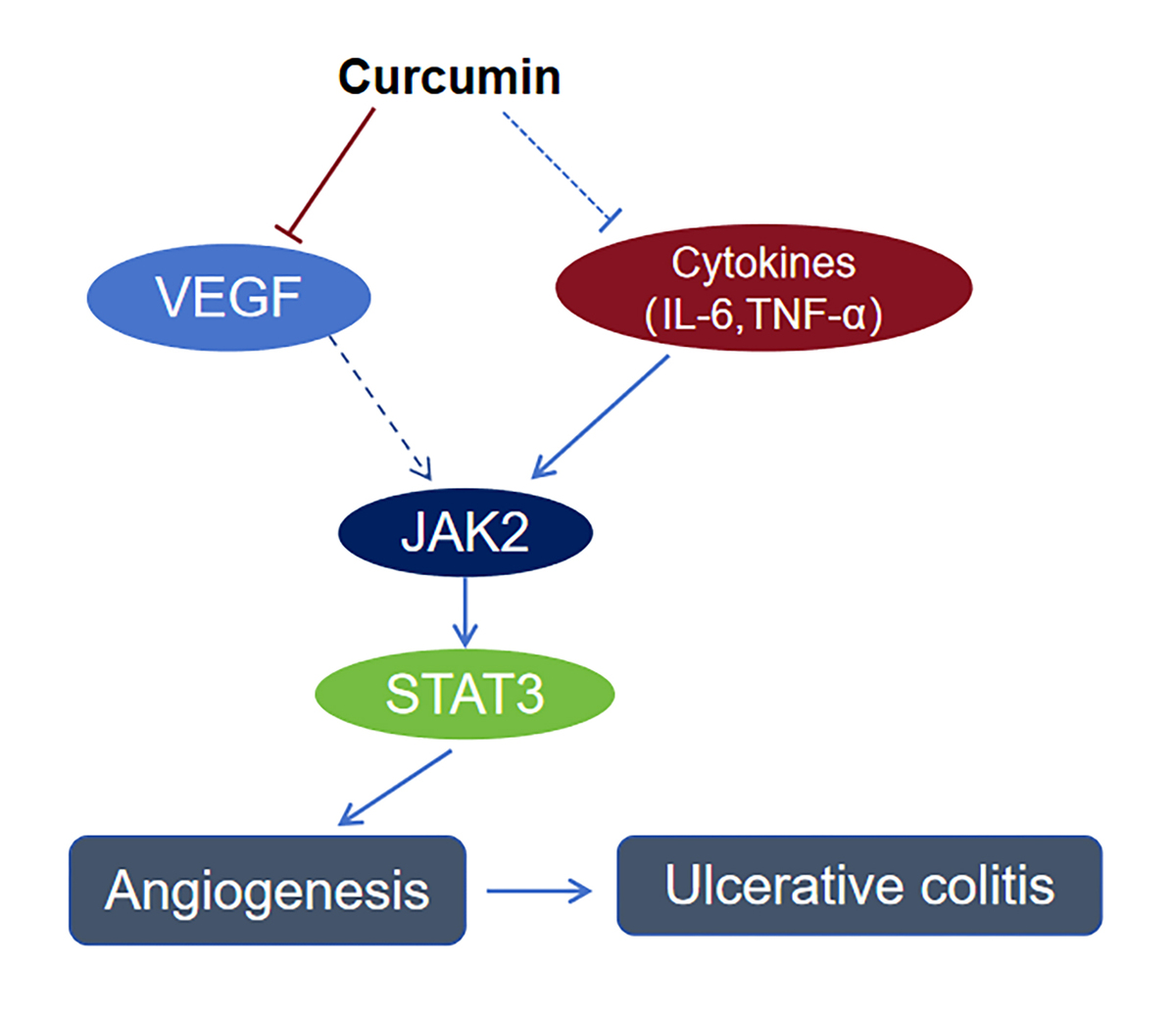
Curcumin, a polyphenolic compound derived from the rhizome of Curcuma longa, exhibits a broad spectrum of pharmacological activities, making it a subject of extensive research in both preclinical and clinical studies [9]. Its most notable bioactivities include antitumor [10], anti-inflammatory [11], and antifibrotic properties [12], underpinned by its ability to modulate diverse molecular pathways. Many articles have witnessed the improved effect of curcumin on inflammatory diseases including colitis. Curcumin blocks nuclear translocation of NF-κB and suppresses downstream cytokines (e.g., TNF-α, IL-1β, IL-6), resulting in improvement of UC [13]. By binding to Toll-like receptor 4 (TLR4), curcumin disrupts TLR4-mediated NF-κB activation and reduces intestinal permeability [14]. Curcumin downregulates JAK2/STAT3 phosphorylation, curtailing Th1/Th17-driven inflammation and promoting Treg cell activity [15, 16]. Curcumin attenuates ulcerative colitis via regulating the sphingosine kinase 1/NF-κB signaling pathway [17]. Furthermore, curcumin attenuates visceral hyperalgesia through inhibiting phosphorylation of TRPV1 in a rat model of ulcerative colitis [18]. Curcumin arrests dendritic cells of maturation to promote Treg function, shifting the balance toward anti-inflammatory responses [19].
Angiogenesis, the formation of new blood vessels from pre-existing ones, is a tightly regulated process critical for both physiological development and pathological conditions such as cancer and chronic inflammation [20]. It involves complex interactions between endothelial cells, signaling molecules, and extracellular matrix components, enabling tissue growth, repair, and nutrient supply [21]. Pathological angiogenesis, however, often exhibits abnormal vessel structures and hyperpermeability, contributing to tumor progression and metastasis [22]. However, mounting evidence indicates a close relationship between inflammatory bowel disease and angiogenesis [23].
Considering the overall evidence, we hypothesized that curcumin may ameliorate ulcerative colitis via inhibiting angiogenesis. Concurrently, we aimed to elucidate its molecular mechanism.
Material and methods
Chemicals and reagents
Curcumin was obtained from Nanjing Jin Yibai Biological Technology Co. Ltd. (Nanjing, China). TNF-α (KE10002) and IL-6 (KE10007) ELISA kits were purchased from Proteintech. The following antibodies were used: VEGF (Proteintech, 66828-1-lg), VEGFR (Proteintech, 83049-4-RR), JAK2 (Abcam, 245303), p-JAK2 (Abcam, 32101), STAT3 (Proteintech, 51076-1-AP) and p-STAT3 (Y705) (Abcam, 278669), GAPDH (Proteintech, 10494-1-AP).
Animals
The male C57 mice (20–25 g) were purchased from the Shanghai SLAC Laboratory Animal Co., Ltd. Every 5 mice were housed as a group in a cage, and provided with free access to food and water at controlled room temperature (22 ±2°C) and humidity (60–80%) under a 12 h/12 h light/dark cycle. The animal experiment was approved by the Yancheng TCM Hospital affiliated with Nanjing University of Chinese Medicine Institutional Animal Care and Use Committee (SLDW231201).
Cell culture
HUVEC cells were obtained from the American Type Culture Collection (ATCC) and subjected to regular testing for mycoplasma contamination. HUVECs were cultured in DMEM/F12 medium supplemented with 10% FBS (Cellmax, Lanzhou, China) and 1% penicillin-streptomycin (Corning, NY, USA). Cells were maintained in a humidified incubator at 37°C with 5% CO2 and routinely screened for mycoplasma contamination.
Tube formation assay
Tube formation assay was performed as described previously [24]. Briefly, HUVECs, pretreated with or without curcumin for 24 h, in the presence or absence of VEGF-1 (10 ng/ml) or 2% FBS, were trypsinized and then seeded into a 96-well plate (2 × 104/well) precoated with 40 µl (10 mg/ml) growth factor-reduced Matrigel (BD Biosciences, Billerica, MA). After 2 to 3 h of incubation at 37°C, the capillary tube structures were observed, and representative images were captured with an inverted phase-contrast microscope (Zeiss, Germany) (200×) equipped with the Quick Imaging system. The formed tubes were quantified by ImageJ software. Briefly, three randomly selected fields of view were photographed in each treatment. Tube formation was assessed by counting the circular structures representing tube-like networks. The average of three fields was taken as the value for each treatment.
Rat aortic ring assay
The rat aortic ring assay was performed following the modified methods as described [25]. Thoracic aortas were excised from Sprague-Dawley male rats and cut into cross sectional, 1-mm-wide rings and flushed with DMEM/F12 medium (Hyclone, USA). Rings were immediately placed into the wells of a 48-well plate containing 400 µl fibrinogen/thrombin solution and then incubated at 37°C until the fibrinogen/thrombin solution polymerized. The wells were then overlaid with 600 µl of DMEM/F12 medium containing 10 ng/ml VEGF and various concentrations of curcumin. On the seventh day, vascular vessels sprouting from each ring were examined using a Leica inverted phase-contrast microscope (Leica, Germany) equipped with the Quick Imaging system to take photos and to facilitate counting.
Western blotting analysis
Proteins were extracted from cells using RIPA lysis buffer. Following separation by SDS-PAGE, the proteins were transferred onto polyvinylidene difluoride (PVDF) membranes. These membranes were then blocked with 5% skim milk and probed with specific antibodies targeting anti-JAK2 (1 : 1000), anti-p-JAK2 (1 : 1000), anti-STAT3 (1 : 1000), anti-p-STAT3 (1 : 1000), and anti-actin (1 : 1000). After washing with Tris-buffered saline with Tween 20 (TBST), the relative protein expression was determined using an imaging system (Bio-Rad, USA).
Dextran sulfate sodium (DSS)-induced ulcerative colitis in mice
A total of 40 male Balb/c mice were acclimated for 1 week, and then randomly divided into 4 groups (n = 10): the blank group, control group, and two curcumin groups (25 mg/kg and 50 mg/kg). All groups except the blank group were treated with water containing 4% DSS for 7 days. Simultaneously, the mice of treated groups were administered curcumin (25 mg/kg and 50 mg/kg), whereas the mice of blank and control group were treated with sterile water. During the experiment, all the mice were treated with a standard diet, and the gavage volume was 0.1 ml/10 g. The weight of mice was recorded every day, and the disease activity index score (DAI score) was assessed according to the procedure described by Hao et al. [26]. At the endpoint of the experiment, the colon was dissected and the length of the colon was measured after the blood was prepared. The colon tissues were fixed in 4% neutral formalin for H&E staining and immunohistochemical analysis.
RNA isolation and quantitative real-time PCR
Total RNA was isolated from cells using Trizol, and cDNA was synthesized using the HiScript III-RT SuperMix kit (Vazyme, R323-01). The cDNA samples were subjected to a real-time quantitative polymerase chain reaction (RT-qPCR) using the ChamQ Universal SYBR qPCR Master Mix (Vazyme, Q711-02) performed on an iQ6 instrument (Bio-Rad, USA). The primer sequences were as follows: VEGF: Forward: 5’-TTGCCTTGCTGCTCTACCTCCA -3’. Reverse: 3’-GATGGCAGTAGCTGCGCTGATA -5’
VEGFR: Forward: 5’-GGAACCTCACTATCCGCAGAGT -3’. Reverse: 3’-CCAAGTTCGTCTTTTCCTGGGC -5’
H&E staining
The harvested lung and liver tissues were first rinsed with PBS and subsequently fixed in 4% paraformaldehyde for over 24 h. After fixation, the tissues underwent dehydration with a gradient of ethanol and xylene before being embedded in paraffin. Sections of 4 µm thickness were then prepared and stained with hematoxylin and eosin (H&E) following standard protocols. Finally, the sections were imaged using a Pathology Workstation (PerkinElmer, USA). Results were quantified using Image J software (Bethesda, NIH, USA).
Immunohistochemical staining
Lung tissue sections collected from mice were first rehydrated and then subjected to antigen extraction in 10 mM sodium citrate and 0.05% Tween solution at 95°C for 3 min. Sections were then blocked with 5% bovine serum albumin (BSA) solution for 30 min, followed by incubation with the indicated primary antibodies overnight at 4°C and then with the corresponding secondary antibodies for 2 h at room temperature. Finally, the sections were stained with 3,3ʹ-diaminobenzidine (DAB) (ZSGB-BIO, ZLI-9018), sealed with neutral gum, and observed under a Pathology Workstation (PerkinElmer, USA). The immunohistochemical signals were photographed and the images were analyzed using ImageJ software (Bethesda, NIH, USA).
Results
Comprehensive target analysis of curcumin, ulcerative colitis, and angiogenesis using multiple databases
Curcumin, ulcerative colitis, and angiogenesis-related targets were collected to investigate potential mechanisms of action. Using the Swiss Target Prediction website, the SMILES sequence of curcumin (PubChem CID: 969516) was input, selecting Homo sapiens as the species. This yielded 65 targets. Using the PharmMapper website, the structural file of curcumin was uploaded, selecting Human Protein Targets Only. Targets were filtered for Norm Fit ≥ 0.5. IDs were then mapped to Gene Names using the UniProt database, resulting in 106 targets. Using the STITCH website, the SMILES sequence of curcumin was input, selecting Homo sapiens and filtering for low confidence (0.150). Exported data yielded 10 targets. The curcumin targets obtained from the above sources were combined and duplicate targets were removed, resulting in a total of 170 targets.
Using the GeneCards website, a search was performed with the keyword of Ulcerative Colitis. The downloaded data contained 5,843 genes. Targets with a score greater than the median score of all genes were screened, resulting in 2,921 targets. Using the GeneCards, OMIM, and TTD websites, the same search was performed with the keyword Angiogenesis. Genes were copied into a table. The angiogenesis-related targets obtained from the above sources were combined and duplicate targets were removed, resulting in 1,723 targets.
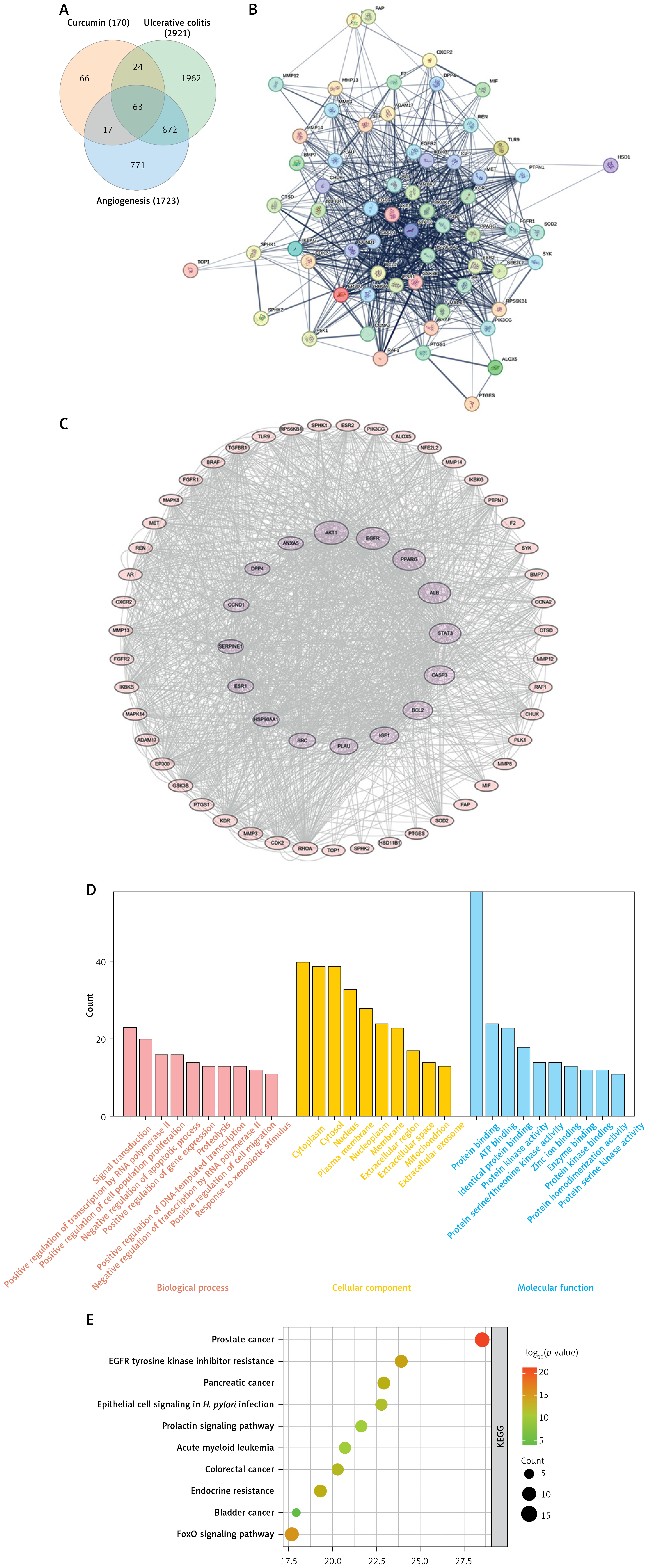
Finally, a Venn diagram was constructed using the obtained target datasets, revealing 63 common intersection targets (Figure 1 A). A protein-protein interaction (PPI) network was constructed using the STRING website. “Multiple proteins” was selected, and the common intersection targets were input. The resulting image and a TSV data file were exported (Figure 1 B). The TSV data were imported into Cytoscape software. Targets were ranked based on their degree value, and the PPI network diagram was visualized (Figure 1 C). The top ten ranked targets were: AKT1, EGFR, PPARG, ALB, STAT3, CASP3, BCL2, IGF1, PLAU, SRC. Using the DAVID database, the common overlapping targets were imported for GO/KEGG enrichment analysis. The GO analysis results were primarily enriched in processes such as signal transduction, transcriptional regulation by RNA polymerase, and cell proliferation (Figure 1 D). KEGG pathway analysis showed significant enrichment in pathways including prostate cancer, EGFR tyrosine kinase inhibitor resistance, pancreatic cancer, epithelial signaling transduction, and acute myeloid leukemia (Figure 1 E). Among the identified targets, STAT3 was chosen for further investigation.
Figure 1
Comprehensive target analysis using multiple databases. Multiple databases were used to comprehensively analyze the possible targets for curcumin, ulcerative colitis, and angiogenesis, then the following diagrams were drawn using the corresponding software in the website: A – Venn diagram, B – Network diagram of targets, C – PPI diagram, D – GO analysis, E – KEGG analysis
Curcumin inhibits tube formation and vascular sprouting in vitro
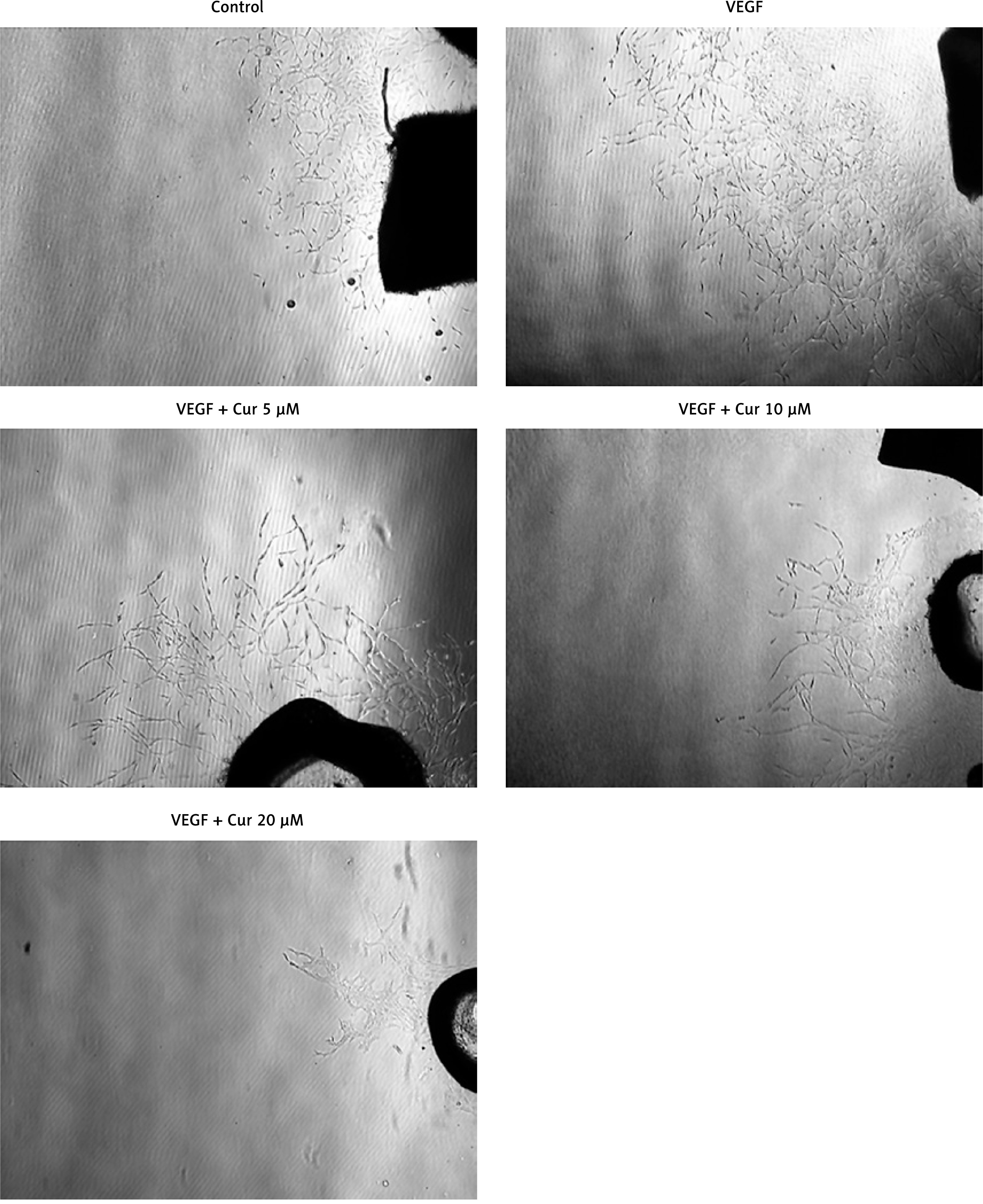
To confirm curcumin’s effect on angiogenesis, tube formation assay and rat aortic ring assay were used. As shown in Figure 2, VEGF clearly induced tube formation, while curcumin significantly inhibited VEGF-induced tube formation in HUVECs in a concentration-dependent manner. Figure 3 shows that the number of sprouting endothelial cells in the curcumin treatment group was significantly lower than that of the control group. Both results suggested that curcumin inhibited angiogenesis in vitro.
Figure 2
Curcumin suppresses tube formation in vitro. HUVECs were treated with curcumin (0–20 μM) in vitro in the presence or absence of VEGF (10 ng/ml) for 24 h, then harvested to be seeded in a 96-well plate precoated with 40 μl (10 mg/ml) growth factor-reduced Matrigel (2 × 104 cells/well). After 2–3 h incubation, the capillary tube structures were observed and the images were captured with an inverted phase-contrast microscope. A – Representative images (200×) of tube formation. B – Tube-like structures were evaluated using ImageJ software. Quantitative data are presented as mean ± SD (n = 3). Comparison with control, ##p < 0.01; comparison with VEGF group, **p < 0.01
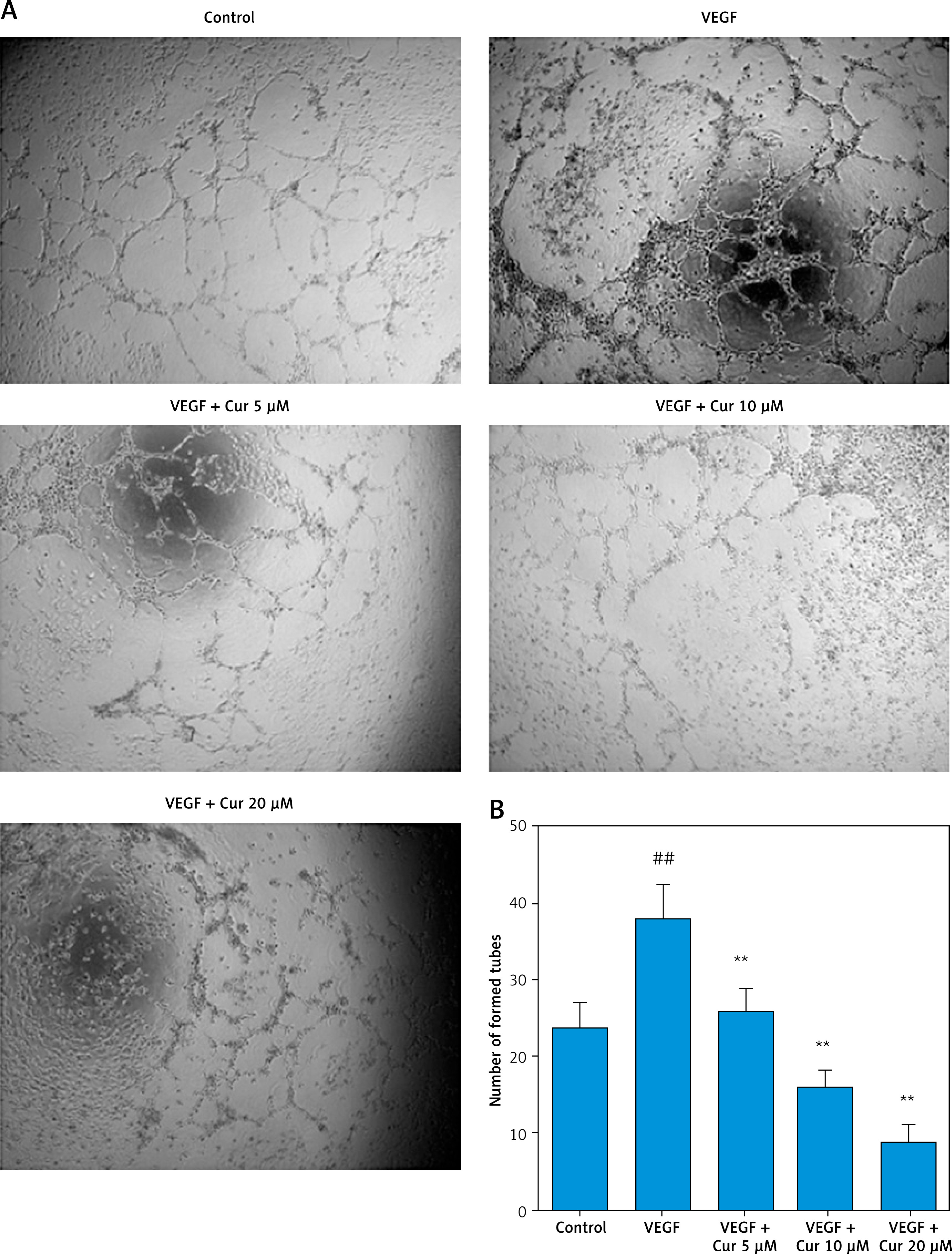
Figure 3
Curcumin inhibits vascular sprouting of rat aortic ring in vitro. Rat aortic rings were placed into the wells of a 48-well plate containing 400 μl of fibrinogen/thrombin solution and then incubated at 37°C for polymerization. The wells were then overlaid with 600 μl of DMEM/F12 medium containing 10 ng/ml VEGF and various concentrations of curcumin. After 6-day incubation, vascular vessels sprouting from the rat aortic ring were photographed at 200× magnification using a Leica inverted phase-contrast microscope equipped with the Quick Imaging system (n = 3)
Curcumin inhibits VEGF expression and the JAK2/STAT3 pathway in HUVEC cells
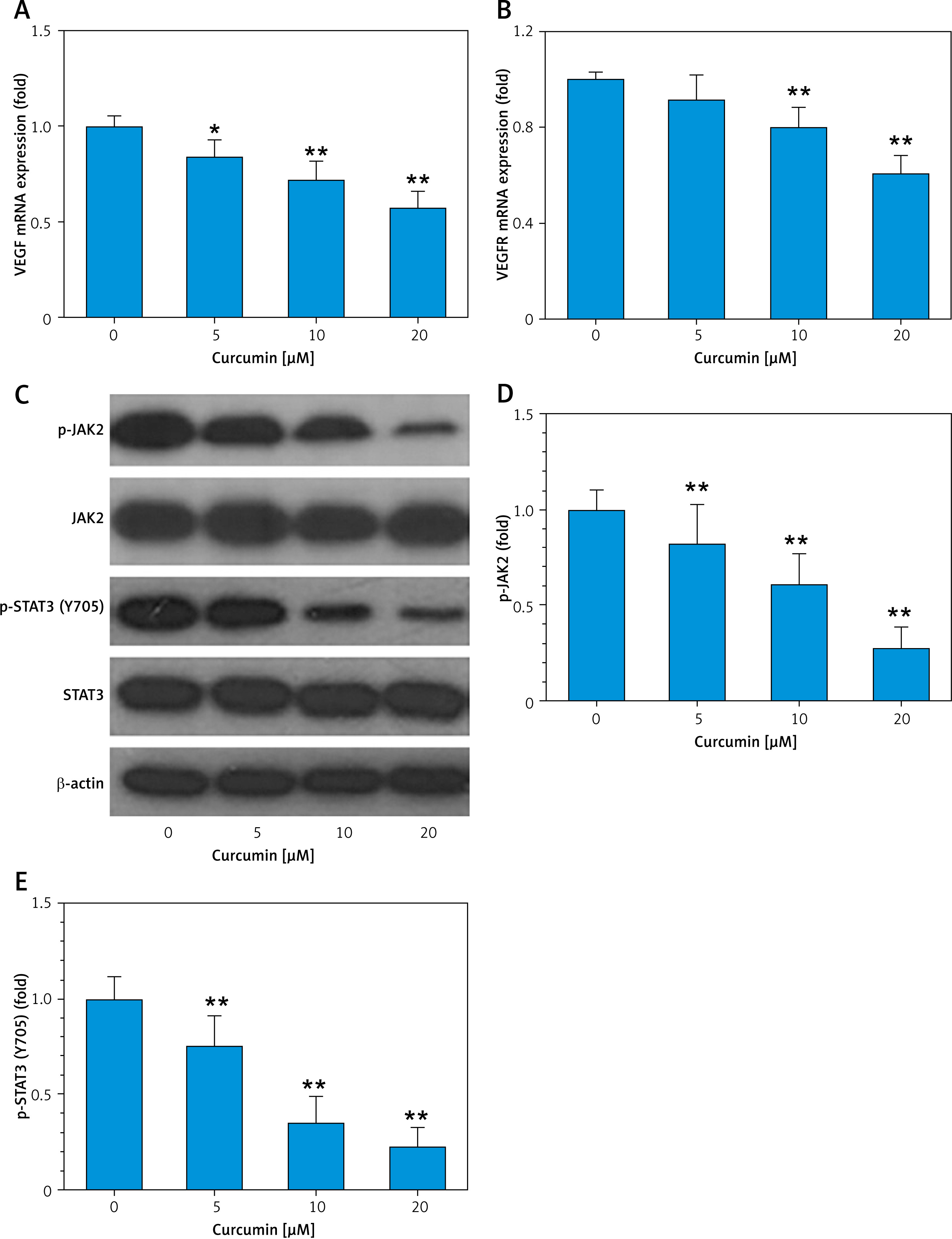
We next observed how curcumin affects HUVEC cells in vitro. RT-PCR results showed that curcumin could inhibit transcriptional expression of VEGF and VEGFR (Figures 4 A, B). Western blotting data demonstrated curcumin’s inhibitory effect on phosphorylation of JAK2 and phosphorylation of STAT3 but not on total JAK2 and STAT3 protein levels (Figures 4 C–E). These results indicated that the VEGF-mediated JAK2/STAT3 pathway was involved in curcumin’s inhibition of angiogenesis.
Figure 4
Curcumin inhibits VEGF and JAK2/STAT3 pathway in HUVEC cells. 5 × 105 cells/well HUVECs were placed in a 6-well plate, and treated with curcumin (0–20 μM) for 8 h. Then cells were harvested for preparation of cellular lysates, and the lysates were subjected to RT-PCR and western blot analysis with antibodies to JAK2 and STAT3. β-actin was used as a control. A – RT-PCR result of VEGF mRNA expression. B – RT-PCR result of VEGFR mRNA expression. C – Western blotting analysis for JAK2/ STAT3. D, E – Quantitative analysis of p-JAK2 and p-STAT3 (Y705). n = 3. Comparison with curcumin 0 μM, *p < 0.05, **p < 0.01
VEGF attenuates curcumin’s effect on the JAK2/STAT3 pathway
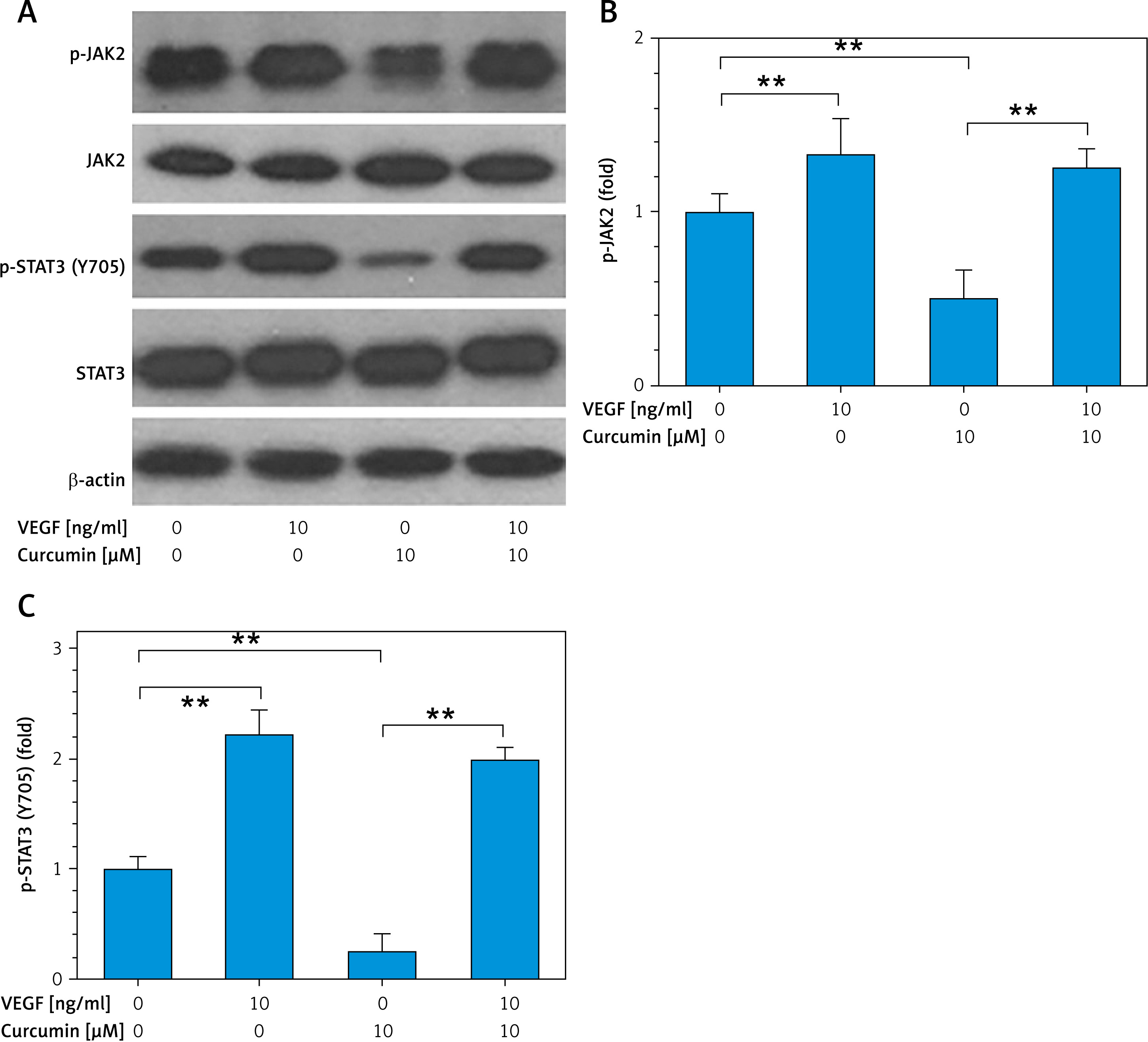
To further confirm whether VEGF mediates curcumin inhibition of the STAT3 pathway, 10 ng/ml VEGF was added to the culture medium of HUVEC cells. Western blotting data showed that VEGF activated p-JAK2 and p-STAT3 in HUVEC cells and curcumin inhibited p-JAK2 and p-STAT3, whereas the inhibitory effect on p-JAK2 and p-STAT3 was significantly attenuated when VEGF was co-administered (Figure 5). This indicated that VEGF mediated the inhibitory effect of curcumin on the JAK2/STAT3 signaling pathway.
Figure 5
Curcumin inhibition of JAK2/STAT3 is partially attenuated by VEGF. 5 × 105 cells/well HUVECs were placed in a 6-well plate, and treated with or without curcumin (10 μM) and VEGF (10 ng/ml) for 8 h. Cells were harvested for preparation of cellular lysates, then the lysates were subjected to western blot analysis with antibodies to JAK2 and STAT3. β-actin was used as a control. A – Western blotting analysis for JAK2/STAT3. B, C – Quantitative analysis of p-JAK2 and p-STAT3 (Y705) by ImageJ software. n = 3. **p < 0.01
Curcumin inhibits DSS-induced colitis in mice through STAT3-mediated angiogenesis
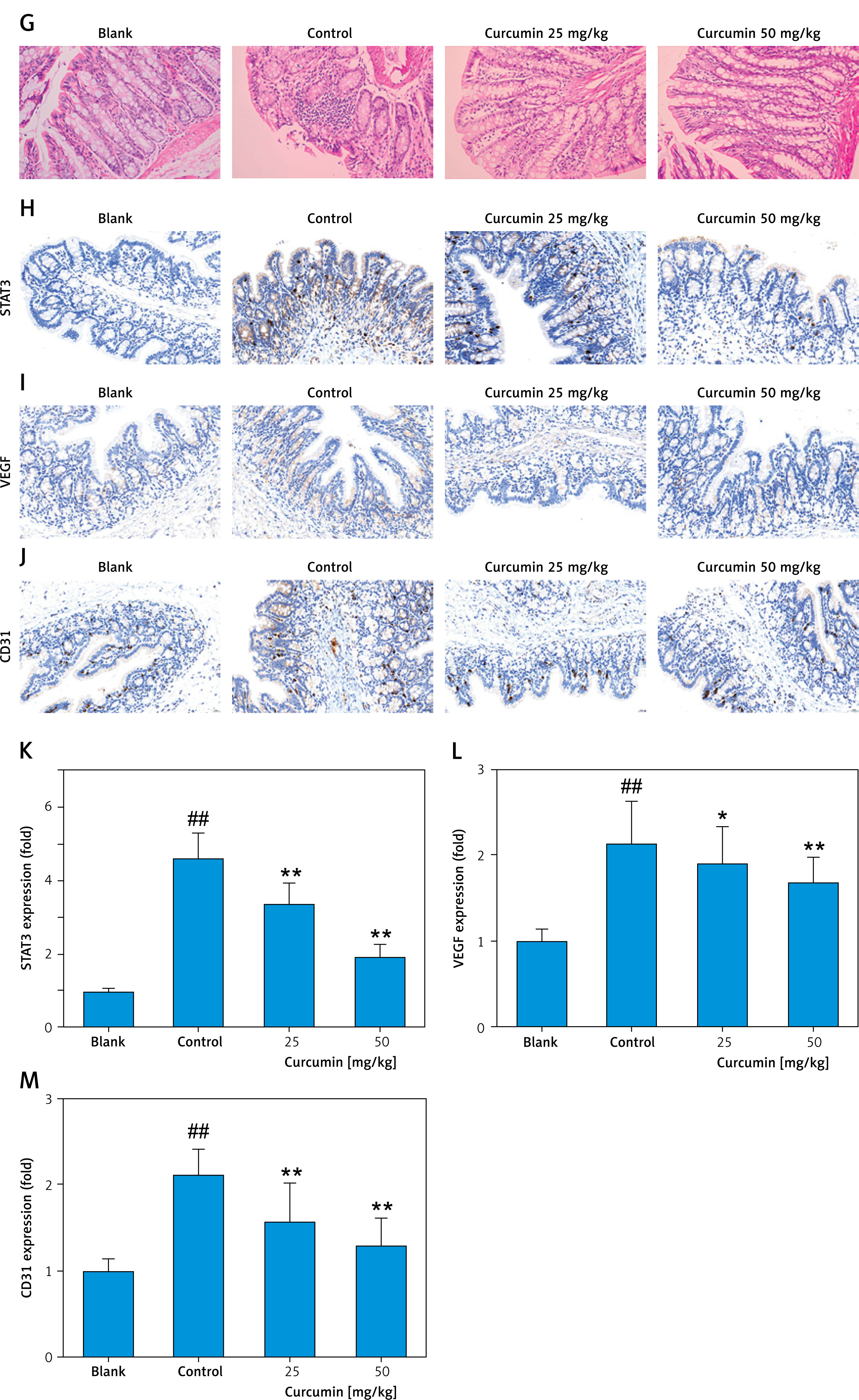
DSS-induced colitis in mice was established according to the experimental flow diagram (Figure 6 A). The DSS-treated mice exhibited marked weight loss, diarrhea, bloody stools, and other colitis symptoms. Compared with the blank group, the length of the colon of the model group was shorter (Figure 6 B), and the DAI scores of weight loss, diarrhea, and bloody stool were significantly higher (Figure 6 C). Compared with the control group, curcumin alleviated colon shortening in DSS-induced colitis mice (Figure 6 B). A similar result was achieved in terms of DAI score; curcumin notably reduced the DAI score by improving the weight loss, diarrhea, and bloody stool of the DSS-induced colitis mice (Figure 6 C). In the serum of mice with ulcerative colitis, TNF-α content was significantly elevated, and curcumin inhibited TNF-α expression in serum (Figure 6 D). Similar results were observed for IL-6 and IL-1β content in serum (Figures 6 E, F). The results of H&E staining indicated that, compared with the blank group, the colon of control group mice exhibited severe mucosal damage, inflammatory cell infiltration, and gland structure damage (Figure 6 G). However, curcumin partially reversed colonic mucosal injury, eliminated inflammatory cell infiltration, and alleviated destruction of glands (Figure 6 G). The further immunohistochemical staining results showed that expression of STAT3 and VEGF was markedly elevated in the tissue of colitis colon, whereas it was decreased in the groups with curcumin treatment (Figures 6 H, I, K, L). Also, CD31 expression analysis suggested that curcumin inhibited colonic angiogenesis in mice (Figures 6 J, M). Overall, the above results suggested that curcumin exerted therapeutic effect on ulcerative colitis and the molecular mechanism was related to inhibition of the VEGF-activated JAK/STAT3 pathway.
Figure 6
STAT3-promoted angiogenesis mediates curcumin inhibition of DSS-induced colitis in mice. To induce ulcerative colitis, the mice were treated with water containing 4% DSS for 7 days. Simultaneously, the mice of treated groups were gavaged with curcumin (25 mg/kg and 50 mg/kg) for 7 days, whereas mice in the blank and control groups received sterile water. During or at the endpoint of the experiment, the following assays were conducted: A – Experimental flow diagram. B – Length of colon. C – DAI score in colon. D – ELISA assay for TNF-α. E – ELISA assay for IL-6. F – ELISA assay for IL-1β. n = 10. Comparison with blank, ##p < 0.01; comparison with control, *p < 0.05, **p < 0.01, G – H&E staining of colon tissues (×200). Immunohistochemical staining of STAT3 (H), VEGF (I), and CD31 (J), and the quantitative analysis of STAT3 (K), VEGF (L), and CD31(M) was assessed (×200). Comparison with blank, ##p < 0.01; comparison with control, *p < 0.05, **p < 0.01
Discussion
According to the hypothesis, we firstly used the tools of multiple databases to analyze the possible targets of curcumin against UC, and the top ten proteins – AKT1, EGFR, PPARG, ALB, STAT3, CASP3, BCL2, IGF1, PLAU, SRC – were screened. Among these proteins, STAT3 is closely involved in inflammation. Combined with curcumin’s potent anti-inflammatory activity [11, 27], STAT3 is regarded as a mediator of angiogenesis regulating UC. Central to angiogenesis regulation is vascular endothelial growth factor (VEGF), a master cytokine that promotes endothelial cell proliferation, migration, and vascular permeability [28]. VEGF-A, the most studied isoform, is upregulated by hypoxia and promotes sprouting angiogenesis by activating receptors such as VEGFR-2, triggering downstream signaling pathways including PI3K/AKT and MAPK [28]. In tumors, VEGF overexpression drives aberrant vascular networks, facilitating metastasis and immune evasion [29].
In UC, chronic inflammation triggers excessive and disorganized blood vessel formation through several interconnected mechanisms [30]. Tissue hypoxia in inflamed mucosa upregulates hypoxia-inducible factor-1α (HIF-1α), leading to VEGF overexpression. VEGF drives immature, hyperpermeable vessel formation [23]. These vessels exhibit poor barrier function, facilitating leukocyte extravasation and edema, and further aggravating mucosal injury [23]. Notably, VEGF is not merely a bystander; its levels correlate with disease severity and internal inflammation [31].
Recent studies highlighted the interplay between VEGF and STAT3, a transcription factor activated by reactive oxygen species (ROS) or cytokine signaling [32]. ROS-mediated STAT3 activation enhances VEGF expression, forming a feedforward loop that sustains angiogenesis [33]. For instance, 27-hydroxycholesterol (27HC), a tumor-promoting metabolite, induces STAT3 phosphorylation via ROS, leading to VEGF autocrine signaling and endothelial recruitment in both estrogen receptor-positive and -negative cancers [33]. Inhibiting STAT3 or ROS effectively suppresses VEGF-driven angiogenesis, underscoring their therapeutic potential [34]. Together, VEGF and STAT3 represent pivotal targets for modulating angiogenesis in diseases, with ongoing research exploring dual-pathway inhibitors to combat pathological vascularization. Thus, targeting pathological angiogenesis offers novel avenues for UC treatment.
The present study demonstrated that curcumin improved DSS-induced UC in mice. Further results showed that the therapeutic effects of curcumin on UC are related to inhibition of angiogenesis. Among the potential mechanisms, inhibition of the VEGF-mediated JAK2/STAT3 pathway was demonstrated to be involved in this process. Curcumin can inhibit several types of cancers [35, 36] and improve acute renal injury [37] and liver injury [38] via inhibition of the JAK2/STAT3 pathway. These findings suggest that JAK2/STAT3 is a key regulatory molecule mediating the pharmacologic activities of curcumin.
Aberrant angiogenesis is not merely a secondary phenomenon but a central pathological pillar in UC, driving ischemia, inflammation, and impaired tissue repair. Targeting specific pathways through localized VEGF inhibition holds transformative potential. Future therapies should aim to normalize vascular function rather than indiscriminately inhibit angiogenesis, thereby promoting true mucosal healing. Combining these vascular-targeted strategies with conventional immunomodulation offers a promising roadmap for overcoming therapeutic resistance in complex UC cases. Furthermore, based on its extensive activity, we inferred that curcumin likely combated UC by synergistically modulating inflammation, oxidative stress, immune responses, and microbiota dysbiosis. Simultaneously targeting multiple proteins related to UC may achieve a better curative effect than a single protein target.